Географическое положение и климат
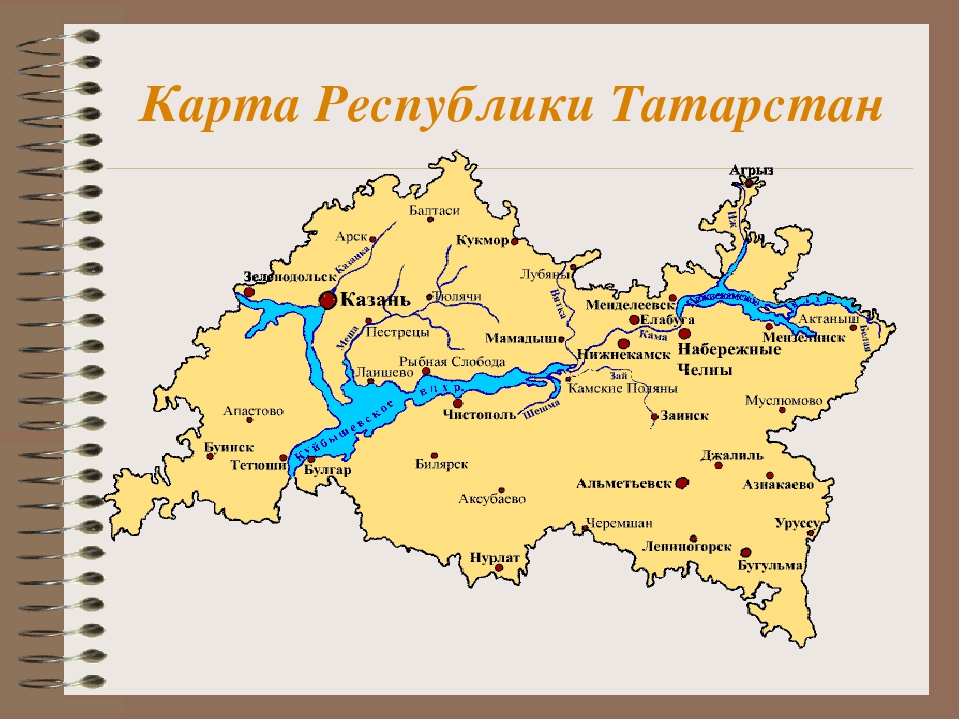
Татарстан расположен на востоке Восточно-Европейской равнины, в месте слияния двух крупнейших рек – Волги и Камы, г.Казань находится на расстоянии 797 км к востоку от г.Москвы.Общая площадь республики составляет 6783,7 тыс.га. Максимальная протяженность территории – 290 км с севера на юг и 460 км с запада на восток. Границ с иностранными государствами Татарстан не имеет.
Территория Татарстана представляет собой возвышенную ступенчатую равнину, расчлененную густой сетью речных долин. Широкими долинами Волги и Камы равнина разделена на три части: Предволжье, Предкамье и Закамье. Предволжье с максимальными высотами 276 м занимает северо-восточную часть Приволжской возвышенности. В Восточное Предкамье с севера заходят южные окончания Можгинской и Сарапульской возвышенностей, разделенные долиной р.Иж. Наибольшие высоты достигают здесь 243 м. Самой высокой в Татарстане (до 381 м) является Бугульминская возвышенность в Восточном Закамье. Самый низкий рельеф (в основном до 200 м) характерен для Западного Закамья.
17% территории республики покрыто лесами, состоящими из деревьев преимущественно лиственных пород (дуб, липа, береза, осина), хвойные породы представлены сосной и елью. На территории Татарстана обитают 433 вида позвоночных, а также несколько тысяч видов беспозвоночных животных.
Территория Татарстана характеризуется умеренно-континентальным типом климата средних широт, с теплым летом и умеренно-холодной зимой. Самым теплым месяцем является июль со средней месячной температурой воздуха по территории 18 – 20 °С, самым холодным – январь со средними месячными температурами от -13 °С. Продолжительность теплого периода (с устойчивой температурой выше 0 °С) колеблется по территории в пределах 198-209 дней, холодного – 156-167 дней. Осадки по территории распределяются сравнительно равномерно, годовая сумма их составляет 460 – 540 мм.
Почвы отличаются большим разнообразием – от серых лесных и подзолистых на севере и западе до различных видов черноземов на юге республики.
На территории Татарстана расположены Волжско-Камский государственный природный биосферный заповедник и национальный парк «Нижняя Кама». Волжско-Камский государственный природный биосферный заповедник расположен на территории Зеленодольского и Лаишевского муниципальных районов Республики Татарстан. Два обособленных участка заповедника — Сараловский (4170 га) и Раифский (5921 га) удалены друг от друга на расстояние около 100 км. Национальный парк «Нижняя Кама» расположен на территории двух муниципальных районов Республики Татарстан: Елабужского и Тукаевского. На территории парка намечено несколько сухопутных и водных туристских маршрутов по лесным массивам, а также водные маршруты по акватории водохранилища, по рекам Каме и Криуше.
Волжско-Камский государственный природный биосферный заповедник расположен на территории Зеленодольского и Лаишевского муниципальных районов Республики Татарстан. Два обособленных участка заповедника — Сараловский (4170 га) и Раифский (5921 га) удалены друг от друга на расстояние около 100 км. Национальный парк «Нижняя Кама» расположен на территории двух муниципальных районов Республики Татарстан: Елабужского и Тукаевского. На территории парка намечено несколько сухопутных и водных туристских маршрутов по лесным массивам, а также водные маршруты по акватории водохранилища, по рекам Каме и Криуше.
Добро пожаловать в Республику Татарстан!
Республика Татарстан
Татарстан расположен в месте слияния двух крупнейших рек Волги и Камы и служит своего рода мостом между европейской и азиатской частью России.
Республика по площади (67,8 тыс.кв.км) больше таких европейских стран как Бельгия или Нидерланды и примерно равна Ирландии.
В Татарстане веками вместе проживают татары (53,2%) и русские (39,7%), чуваши и удмурты, мордва и марийцы, украинцы и башкиры — всего 173 национальности, 3,8 млн. человек.
Татарстан — динамично развивающийся регион, успехи которого базируются на прочной основе экономической стабильности. Республика известна в России и за рубежом как центр нефтедобычи и нефтехимии, производитель всемирно известных грузовиков КАМАЗ и вертолетов МИ, один из лидеров российского сельскохозяйственного производства.
Являясь местом встречи цивилизаций, символическим «перекрестком» религиозных и культурных традиций Востока и Запада, Азии и Европы, Татарстан накопил богатейший опыт межэтнического и межконфессионального согласия. Опираясь на него, республика развивает свои контакты с миром, выстраивает торговые, научные и культурные связи.
Казань — столица Татарстана |
|
|
Столица республики — город Казань с населением свыше 1,1 млн. В преддверии проведения в Казани крупных международных спортивных мероприятий — XXVII Всемирной летней Универсиады 2013 г., Чемпионата мира по водным видам спорта 2015 г. и матчей Чемпионата мира по футболу 2018 г. активно развивается инфраструктура города. |
|
Татарстан — исторический регион |
|
|
Расположенный на пересечении торговых путей Татарстан исторически был призван выполнять роль политического и торгово-экономического центра. Душой и сердцем тысячелетнего города является Казанский Кремль, включенный в список «Всемирного наследия» ЮНЕСКО. Болгар, столица раннефеодального государства Волжской Булгарии, овеян легендами и связан с принятием ислама булгарами — предками казанских татар, расцветом государства Волжской Булгарии в XIII-XIV вв. В живописном устье реки Свияги расположен остров-град Свияжск — уникальный исторический и архитектурно-художественный комплекс второй половины XVI-XX века. После основания в 1551 году города и монастырей он несколько веков был общероссийской православной святыней. Кроме Казани, свой тысячелетний юбилей отметила и Елабуга, город уникальной купеческой архитектуры XIX в., сохранивший до наших дней свой неповторимый облик и уникальный ландшафт. |
|

Культура |
|
|
Знаковыми мероприятиями культурной жизни республики являются крупные международные фестивали: оперный им. В Татарстане свыше сотни музейных учреждений. Посещаемость постоянных экспозиций и выставок музеев превышает 1,7 млн. человек. |
|
 Ф.И.Шаляпина, фестиваль классического балета им. Р.Нуриева, фестиваль современной музыки «Европа-Азия» и фестиваль татарской песни имени Рашита Вагапова. Ежегодно осенью в Казани проходит Международный фестиваль мусульманского кино. С 2008 г. ежегодно в Казани проходит музыкальный фестиваль «Сотворение мира».
Ф.И.Шаляпина, фестиваль классического балета им. Р.Нуриева, фестиваль современной музыки «Европа-Азия» и фестиваль татарской песни имени Рашита Вагапова. Ежегодно осенью в Казани проходит Международный фестиваль мусульманского кино. С 2008 г. ежегодно в Казани проходит музыкальный фестиваль «Сотворение мира».Религии |
|
|
Традиционными для республики конфессиями являются ислам и православие. В Татарстане также представлены католицизм, протестантство, иудаизм и другие конфессии. Соблюдение баланса интересов двух крупных конфессий и равенства всех религий перед законом лежит в основе межконфессионального согласия в республике. В Татарстане реализуется масштабный проект по одновременному восстановлению памятников истории и культуры, религиозных святынь — древнего города Болгар и острова-града Свияжск. Панорама Казанского Кремля, где мечеть Кул Шариф и Благовещенский Собор обрамлены древними крепостными стенами, стала своеобразным символом Татарстана, узнаваемым во всем мире. |
|

Спорт |
|
|
Развитие спорта, как профессионального, так и массового, является приоритетным направлением в республике. Команды Татарстана по разным видам спорта неоднократно завоевывали титулы чемпионов России, выигрывали кубки страны, добивались высоких результатов на международных состязаниях. Достижения футбольного клуба «Рубин», хоккейного клуба «Ак Барс», мужской команды «Синтез» по водному поло, волейбольной команды «Зенит-Казань», команды мастеров по хоккею на траве «Динамо-Казань», баскетбольного клуба «УНИКС» привели к признанию Казани спортивной столицей России. На олимпиаде в Пекине Татарстан был представлен 12 спортсменами, которые завоевали 5 медалей, в том числе — 3 золотые. |
|

Использованы фотографии с сайтов: http://www.kzn.ru, http://gokazan.ru, http://yanarysh.tatarstan.ru/ и http://www.photokzn.ru, а также фото Андрея Шлыкова.
Посещая Татарстан, не забудьте:
- Прогуляться по Казанскому Кремлю и посетить Благовещенский собор и мечеть Кул Шариф.
- Попробовать национальную кухню: треугольник, кыстыбый, чак-чак.
- Посетить достопримечательности Татарстана: древний город Болгар, остров-град Свияжск, Раифский монастырь, Елабугу.
- Принять участие в татарском национальном празднике Сабантуй или в русском фольклорном празднике Каравон,

- Посетить музеи Казани (Национальный музей РТ, Центр «Эрмитаж-Казань», Музей изобразительных искусств, Национальную художественную галерею).
- Увидеть «Ватиканский» список Казанской иконы Божией Матери в Крестовоздвиженском храме,
осмотреть казанские мечети дореволюционной постройки. - Прослушать концерт в Государственном Большом концертном зале, Татарской государственной филармонии, посетить представление в одном из театров столицы Татарстана.
- Приобрести сувенир, например, национальный головной убор – тюбетейку.
- Совершить прогулку по реке Волге, посетить казанский аквапарк.
- Выучить несколько татарских слов.

О республике — Татцентр.ру
Географическое положениеТатарстан расположен в центре Российской Федерации на Восточно-Европейской равнине, в месте слияния двух крупнейших рек Европы — Волги и Камы. Общая площадь республики составляет 67 836,2 тыс. кв. км (0,4% общей площади Российской Федерации). Численность населения в республике составляет 3 млн. 898 тыс. человек. Казань — столица Татарстана — находится на расстоянии 797 км к востоку от Москвы.
Общая площадь республики составляет 67 836,2 тыс. кв. км (0,4% общей площади Российской Федерации). Численность населения в республике составляет 3 млн. 898 тыс. человек. Казань — столица Татарстана — находится на расстоянии 797 км к востоку от Москвы.
Климат — умеренно-континентальный. Время — московское (GMT+04:00). Границ с иностранными государствами Татарстан не имеет. Столица республики — город Казань с населением более 1 млн. 231 тыс. человек. Территория Казани составляет площадь 425,3 кв. км. В Татарстане 43 муниципальных района и 2 городских округа.
Фото С. Козловского
Территория республики представляет собой равнину в лесной и лесостепной зоне с небольшими возвышенностями на правобережье Волги и юго-востоке республики. 90% территории лежит на высоте не более 200 м над уровнем моря.
География РТНа территории Татарстана расположены национальный парк Нижняя Кама, Волжско-Камский заповедник. Волжско-Камский заповедник расположен в Прикамье, на левобережных террасах Волги, на территории Зеленодольского и Лаишевского районов Республики Татарстан. Два обособленных участка заповедника — Сараловский (4170 га) и Раифский (5921 га) удалены друг от друга на 100 км.
Два обособленных участка заповедника — Сараловский (4170 га) и Раифский (5921 га) удалены друг от друга на 100 км.
Национальный парк «Нижняя Кама» расположен на территории двух административных районов Республики Татарстан: Елабужского и Тукаевского. На территории парка намечено несколько сухопутных и водных туристских маршрутов по лесным массивам, а также водные маршруты по акватории водохранилища, по рекам Каме и Криуше.
НаселениеВ Татарстане проживает 3 млн. 893 тыс. 756 человек. Республика Татарстан занимает восьмое место в России по численности населения после городов Москва и Санкт-Петербург, Краснодарского края, Республики Башкортостан, Московской, Свердловской и Ростовской областей. В Приволжском федеральном округе республика — вторая по численности населения.
По сравнению с 01.01.2017 численность населения увеличилась на 8,6 тыс. человек, или на 0,2%. В Татарстане доля городского населения на 01.01.2018 составила 76,8%. Лидирует по числу жителей столица республики г. Казань.
Казань.
Татарстан является одной из самых многонациональных территорий России. По данным Всероссийской переписи населения 2010 года представители свыше 173 национальностей проживают на территории республики, в том числе 8 национальностей, численность населения которых превышала 10 тысяч человек: татары, русские, чуваши, удмурты, мордва, марийцы, украинцы и башкиры. Среди народов, населяющих Татарстан, преобладающие по численности населения — татары (более 2 млн. чел. или 53,2% от общей численности населения республики). На втором месте русские — более 1,5 млн чел. или 39,7%, на третьем — чуваши (116,2 тыс. чел. или 3,1%).
Государственный флаг Республики ТатарстанГосударственный флаг Республики Татарстан представляет собой прямоугольное полотнище с горизонтальными полосами зеленого, белого и красного цветов. Белая полоса составляет 1/15 ширины флага и расположена между равными по ширине полосами зеленого (кобальт зеленый светлый) и красного (кадмий красный светлый) цветов. Зеленая полоса наверху.
Зеленая полоса наверху.
Отношение ширины флага к его длине — 1:2.
Автором Государственного флага Республики Татарстан является Т.Г. Хазиахметов.
Государственный герб Республики ТатарстанГосударственный герб Республики Татарстан представляет собой изображение крылатого барса с круглым щитом на боку, с приподнятой правой передней лапой на фоне диска солнца, помещенного в обрамление из татарского народного орнамента, в основании которого надпись «Татарстан», крылья состоят из семи перьев, розетка на щите состоит из восьми лепестков.
В цветном изображении Государственного герба Республики Татарстан солнце — красного (кадмий красный светлый), барс, его крылья и розетка на щите — белого, обрамление — зеленого (кобальт зеленый светлый), щит, орнамент на обрамлении и надпись «Татарстан» — золотистого цвета.
Авторами Государственного герба Республики Татарстан являются Н.Г. Ханзафаров (идея), Р.З. Фахрутдинов (исполнение).
О Казани — Официальный портал Казани
Казань – столица Республики Татарстан, древний город с многовековой историей, расположенный на левом берегу реки Волга.
Существует несколько легенд, связанных с названием города. По самой распространенной из них Казань происходит от татарского слова «казан» — котел. Легенда гласит, что город был заложен на том месте, где без огня закипела вода в котле, врытом в землю. Символом города, изображенным на гербе, является мифическое драконоподобное существо Зилант.
Казань была основана как форпост на северных границах Волжской Булгарии более 1000 лет назад. Возраст города был определен во время раскопок на территории Казанского Кремля, когда была найдена чешская монета, датированная 929-930 годами, а также остатки каменной кладки и деревянной городской ограды, предметы утвари.
Между Востоком и Западом
Казань – один из ярких примеров того, как в одном городе мирно, в атмосфере дружелюбия и терпимости живут люди разных национальностей и вероисповеданий. Этническое и религиозное разнообразие в столице Татарстана сохраняется не один век.
В столице Татарстана проживают представители свыше 115 национальностей. Самые многочисленные из них по переписи населения 2010 года – русские (48,6%) и татары (47,6%). Также среди жителей Казани можно встретить чувашей, украинцев, марийцев, башкир и удмуртов.
Традиции Казанского университета
Казань – молодежный город, где сосредоточен научно-образовательный центр. В 2019 году город встретил участников мирового чемпионата по профессиональному мастерству «WorldSkills».
Возможность получения доступного и качественного высшего образования привлекает в столицу Татарстана иногородних и иностранных абитуриентов.
Будущие студенты могут выбрать любой из 29 вузов (13 государственных, 9 коммерческих, 7 филиалов вузов), также к их услугам – 34 профессиональных образовательных учреждения. Одним из старейших вузов страны является Казанский (Приволжский) федеральный университет, основанный в 1804 году и славящийся своей научной школой.
Одним из старейших вузов страны является Казанский (Приволжский) федеральный университет, основанный в 1804 году и славящийся своей научной школой.
От Шаляпина до наших дней
Казань имеет уникальный культурный код. На карте города – более 1000 объектов, формирующих культурное многообразие столицы.
В городе открыты двери 34 музеев, 9 театров, 8 крупных концертных залов, 14 дворцов культуры, 50 библиотек и 50 детских школ искусств. В Казани постоянно формируются новые культурные площадки, например, креативное пространство «Штаб», Центр современного искусства «Смена», творческая лаборатория «Угол».
Казань известна на всю страну международными форумами: оперным фестивалем им.Ф.И.Шаляпина, фестивалем классического балета им.Р.Нуриева, фестивалем мусульманского кино, театральным фестивалем тюркских народов «Науруз». Ведущие театры России и звезды мировой величины традиционно включают Казань в свой маршрут гастролей, а музеи города периодически выставляют экспонаты Эрмитажа, Третьяковской галереи и Русского музея.
В 2019 году Казань вступила в сеть креативных городов ЮНЕСКО по направлению «музыка».
Наследие Универсиады
Казань признана спортивной столицей России. «Золотой» страницей в историю города вошла XXVII Всемирная летняя универсиада 2013 года, наследие которой позволило провести XVI чемпионат мира FINA по водным видам спорта 2015 года и матчи Кубка конфедераций FIFA в 2017 году, матчи чемпионата мира по футболу в 2018 году.
Спортивная инфраструктура города включает 15 стадионов, 12 крытых ледовых арен, 54 плавательных бассейна, 378 спортивных зала и 942 плоскостные спортивные площадки.
Настоящим украшением города является 45-тысячный футбольный стадион «Казань Арена» (с 2019 года переименован в «Ак Барс Банк Арену») – первый в России стадион, построенный для проведения мундиаля 2018 года. В 2013 году на «Казань Арене» прошли церемонии открытия и закрытия Всемирных летних студенческих игр. В 2015 году на стадионе состоялись соревнования по плаванию и синхронному плаванию в рамках чемпионата мира по водным видам спорта. В 2017 году «Казань Арена» приняла 4 матча Кубка конфедераций, в 2018 году – шесть матчей чемпионата мира по футболу FIFA.
В 2017 году «Казань Арена» приняла 4 матча Кубка конфедераций, в 2018 году – шесть матчей чемпионата мира по футболу FIFA.
На стадионе свои домашние матчи проводит казанский футбольный клуб «Рубин». Здесь также проходили игры сборной России, матчи Еврокубков.
В приоритете – общественный транспорт
Транспортная сеть Казани по признанию российских и зарубежных экспертов является одной из самых современных и удобных в России. Приоритет в городе отдан общественному транспорту, при этом столица Татарстана первой в России ввела на дорогах выделенную полосу для автобусов и троллейбусов. Сейчас Казань занимает первое место в неофициальном рейтинге городов России по длине выделенных полос для общественного транспорта. В столице Татарстана на 1000 человек приходится 155,7 м выделенных полос.
По результатам исследования Научно-исследовательского института автомобильного транспорта (НИИАТ), который занимается разработкой новой системы транспортного обслуживания Казани, столица Татарстана заняла первое место по показателям эффективности использования маршрутной сети. По улицам Казани курсируют маршруты 4 видов пассажирского транспорта: автобусов, трамваев, троллейбусов и метро. В будние дни общественный транспорт перевозит более 820 тысяч пассажиров.
По улицам Казани курсируют маршруты 4 видов пассажирского транспорта: автобусов, трамваев, троллейбусов и метро. В будние дни общественный транспорт перевозит более 820 тысяч пассажиров.
Миллионы туристов в год
Восхождение на туристический олимп Казань начала в 2005 году, когда столица Татарстана праздновала свое тысячелетие. В юбилейный год город посетило 500 тыс. гостей, в год Универсиады Казань приняла 1,5 млн гостей, в год проведения чемпионата мира по водным видам спорта – 2,1 млн, в 2016 году – 2,5 млн, а в год проведения чемпионата мира по футболу столица Татарстана приняла трехмиллионного туриста. К 2030 году Казань готовится встретить 5-миллионного туриста.
Самыми посещаемыми местами у туристов являются Казанский Кремль, улица Баумана, Центр семьи «Казан», аквапарк «Ривьера» и другие достопримечательности.
В Казани на государственной охране в качестве объектов культурного наследия (памятников истории и культуры) находятся 563 объекта, из них 119 федерального значения, 376 республиканского и 68 местного значений. В 2000 году перечень всемирного культурного наследия ЮНЕСКО пополнился ансамблем Казанского Кремля.
В 2000 году перечень всемирного культурного наследия ЮНЕСКО пополнился ансамблем Казанского Кремля.
Чтобы сохранить рядовую историческую застройку, в окружении которой находятся памятники, Казань включили в список исторических поселений. Более 400 объектов подлежат сохранению без изменения стилистических особенностей. Таким образом, уникальное историческое наследие Казани сохранится для будущих ее поколений.
«Легкие» города
Паркам и скверам в Казани уделяется особое внимание. За последние 9 лет, с 2011 года, при поддержке республики и федерального центра в Казани обновлено и создано с нуля 92 места отдыха. Общая площадь парковых зон увеличилась на 280 га. Сегодня в Казани 144 парка и сквера общей площадью 412 га. Особенно жителям полюбились Горкинско-Ометьевский лесопарк, парк Горького, парк «Черное озеро», парк «Сосновая роща» и парк Победы.
Парки и скверы в Казани – это не только тихое место для отдыха, но и площадки для активного культурного и спортивного досуга горожан.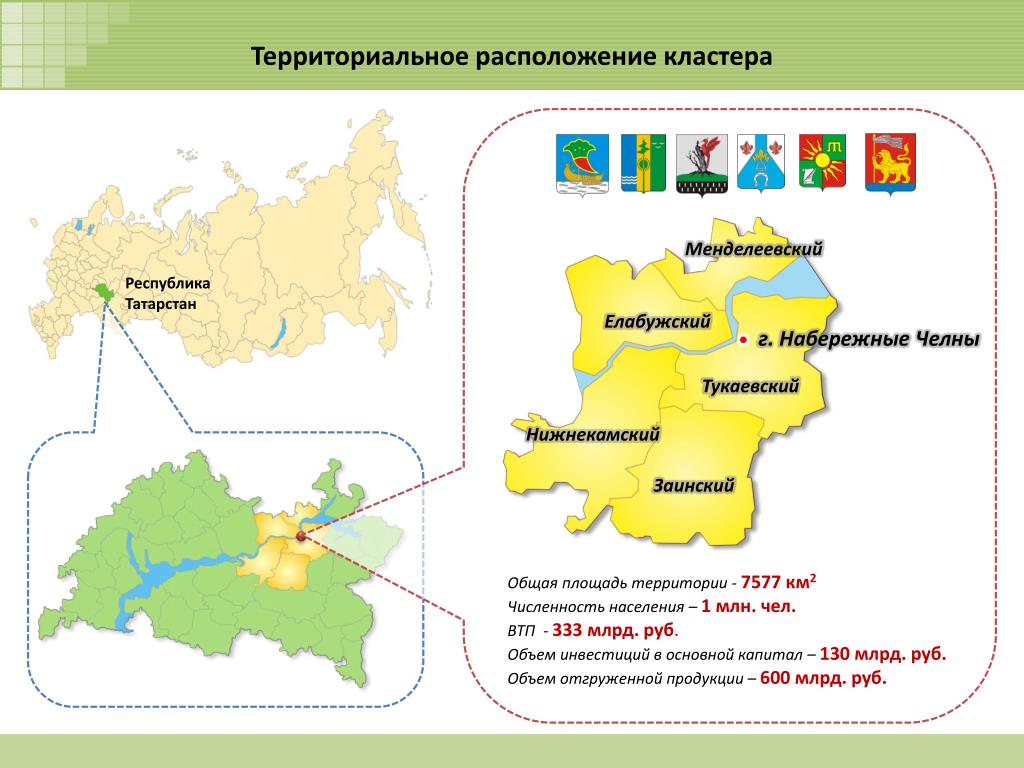 Фестивали, кинопоказы, концерты, благотворительные ярмарки вдохнули жизнь в обновленные общественные пространства.
Фестивали, кинопоказы, концерты, благотворительные ярмарки вдохнули жизнь в обновленные общественные пространства.
Тренд высоких технологий
Отчетливо видно лидерство Казани в сфере экономики. Столица Татарстана занимает передовые позиции по инвестициям и строительству в Поволжье, по обеспеченности современными торговыми центрами и по совокупному капиталу собственных банков в России. В городе расположен крупнейший в России технопарк в сфере высоких технологий «ИТ-парк», а также один из самых больших в Европе технопарков — «Идея».
С 2007 года Мэрия Казани реализует крупномасштабную программу развития малого предпринимательства. Для бизнесменов действуют льготные программы кредитования и аренды муниципального имущества.
Территория и климат
География
Татарстан расположен в центре Европейской части России на Восточно-Европейской равнине, в месте слияния двух крупнейших рек — Волги и Камы.
Столица Республики Татарстан, г.Казань находится на расстоянии 797 км к востоку от Москвы.
Республика Татарстан граничит с восемью регионами Российской Федерации: Республикой Башкортостан, Удмуртской Республикой, Республикой Марий Эл, Чувашской Республикой, Кировской, Оренбургской, Ульяновской и Самарской областями.
Территория республики представляет собой равнину в лесной и лесостепной зоне с небольшими возвышенностями. Около 20 % территории республики покрыто лесами, состоящими из деревьев лиственных и хвойных пород. Татарстан богат водными ресурсами: здесь находится крупных 4 водохранилища, 500 малых рек и более 8000 озер и прудов.
Климат
Территория Татарстана характеризуется умеренно-континентальным типом климата средних широт, с теплым летом и умеренно-холодной зимой. Самый тёплый месяц года — июль, самый холодный — январь.
Лютые морозы и изнуряющая жара не характерны для Татарстана, все 4 сезона года очень хорошо выражены.
За последние 10 лет были отмечены изменения климата в сторону потепления и повышения влажности. Зима в Татарстане становится близкой к прибалтийской, а лето — к средиземноморскому.
Влияние климата на деловую активность
Климат Татарстана влияет на сферу строительства. В основном наружные строительные работы проводятся за 7 месяцев весенне-летнего периода с марта по сентябрь, в остальные 5 месяцев с октября по февраль проходит осенне-зимний сезон, в который проводятся внутренние работы.
Судоходный сезон в Татарстане длится 5 месяцев с мая по сентябрь. Республику пересекает крупнейшая речная магистраль Европы — Волга, связывающая Балтийское, Белое, Каспийское, Азовское и Чёрное моря. Водными путями до Казани можно добраться из Москвы, Санкт-Петербурга, Уфы, Астрахани и других городов.
Мы эффективно используем выгодное географическое положение и климат Татарстана.
Совет Федерации Федерального Собрания Российской Федерации
Флаг и герб региона
ОБЩИЕ СВЕДЕНИЯ
Дата
образования – образована в 1920 году как
Татарская АССР,
в 1991 году –
Татарская ССР,
с 1992 года –
Республика Татарстан (Татарстан).
Административный центр – г. Казань.
Республика Татарстан находится в пределах Приволжского федерального округа
Площадь территории (тыс. кв. км.) – 67,8
Население (тыс. чел.) – 3894,5 (2021 г.)
Национальный состав населения
(по данным Всероссийской переписи населения 2010 г., %):
татары – 53,24;
русские – 39,71;
чуваши – 3,08;
другие национальности – 3,97;
Административно-территориальное устройство (2021 г.):
муниципальные районы – 43
городские округа – 2
городские поселения – 39
сельские поселения – 872
Географическое положение и природно-климатические условия
Республика расположена на востоке Восточно-Европейской равнины, в междуречье Волги и Камы, на стыке центральной России и Урало-Поволжья. Около 90% территории занимают низменные равнины, на западе и юго-востоке – возвышенности: Приволжская и Бугульмино-Белебеевская.
Граничит с Республиками Марий Эл, Башкортостан, Удмуртской и Чувашской республиками, Кировской, Оренбургской, Самарской и Ульяновской областями.
В республике около трех тысяч рек и несколько тысяч крупных и мелких озер. Наиболее крупные водохранилища – Куйбышевское и Нижнекамское.
Главные реки – Волга, Кама, их притоки Вятка, Белая, Свияга, Ик, Иж, Меша, Шешма.
Климат умеренно континентальный. Средняя температура января – минус 11,9°C, июля – плюс 19,2°C.
На территории республики расположены «Волжско-Камский» государственный природный заповедник, национальный природный парк «Нижняя Кама» и ряд государственных природных заказников регионального значения.
Система органов государственной власти
Законодательная власть в Республике Татарстан осуществляется Государственным Советом Республики Татарстан – парламентом Республики Татарстан, который является постоянно действующим высшим представительным, законодательным органом государственной власти республики. Избирается на пять лет.
Государственный Совет Республики Татарстан состоит из 100 депутатов, из которых 50 избираются по одномандатным избирательным округам и 50 – по единому республиканскому избирательному округу по республиканским спискам кандидатов в депутаты, выдвинутым политическими партиями. Действующий состав Государственного Совета Республики Татарстан избран в сентябре 2019 года. Срок окончания полномочий – сентябрь 2024 года.
Исполнительную власть в Республике Татарстан осуществляет Кабинет Министров Республики Татарстан – Правительство Республики Татарстан, являющийся исполнительным и распорядительным органом государственной власти республики. Кабинет Министров Республики Татарстан ответствен перед Президентом Республики Татарстан.
Президент Республики Татарстан является главой государства, высшим должностным лицом Республики Татарстан, срок полномочий – пять лет. Срок окончания полномочий действующего президента республики – сентябрь 2025 года.
Экономика и природные ресурсы
Татарстан является одним из наиболее экономически развитых регионов России.
Среди природных ресурсов республики выделяется нефть. Вместе с нефтью добывается попутный газ, имеются большие запасы нефтяных битумов. Выявлены более 100 залежей угля, привязанные к Южно-Татарскому, Мелекесскому и Северо-Татарскому районам.
Основной вклад в рост валового регионального продукта вносят промышленность, сельское хозяйство и строительство.
Экономика республики в значительной степени ориентирована на экспорт. Ее основу составляет добыча и переработка нефти. Нефтегазохимический комплекс обеспечивает около 50% объема промышленной продукции и является базовым с точки зрения социально-экономического роста, занятости населения и наполнения бюджета всех уровней.
В число крупнейших промышленных предприятий Республики Татарстан входят следующие республиканские предприятия: ПАО «Татнефть», комплекс нефтеперерабатывающих и нефтехимических заводов АО «ТАНЕКО», ПАО «КАМАЗ», ПАО «Казаньоргсинтез».
Наряду с этим основу промышленного потенциала республики составляют предприятия обрабатывающих производств, которые представлены производством машин и оборудования, транспортных средств и оборудования, неметаллических минеральных продуктов, резиновых и пластмассовых изделий, электрооборудования, электронного и оптического оборудования, обработкой древесины и производством изделий из дерева, целлюлозно-бумажным производством, производством нефтепродуктов, легкой и пищевой промышленностью.
Отмечается рост в обрабатывающих производствах, обусловленный в первую очередь восстановлением положительной динамики в производстве транспортных средств и электрооборудования, а также сохранением поступательного тренда в химии и нефтехимии, пищевой промышленности.
В регионе на разных стадиях реализации находятся более сотни инвестиционных проектов. Наибольшая доля в структуре инвестиций приходится на отрасли энергетики и нефтегазохимии.
Наиболее значимым инвестиционным проектом, реализуемым в настоящее время в регионе, является строительство нефтеперерабатывающего комплекса АО «ТАНЕКО» в г. Нижнекамске. Инвестором является ПАО «Татнефть».
Татарстан является одним из лидеров в сфере IT.
Энергетический комплекс представлен тепловыми электростанциями, гидроэлектростанциями и теплоэлектроцентралями.
Агропромышленный комплекс республики практически полностью обеспечивает население основными видами сельхозпродукции. Развито животноводство мясомолочного направления, птицеводство и растениеводство, специализирующееся на выращивании зерна, картофеля, овощей.
Культура и туризм
Культура Татарстана развивалась на стыке двух крупных цивилизаций – восточной и западной, что во многом объясняет ее многообразие.
С Татарстаном связаны судьбы многих выдающихся деятелей культуры: певца Фёдора Шаляпина, писателей Льва Толстого, Сергея Аксакова и Максима Горького, Василия Аксёнова, поэтов Евгения Баратынского, Гавриила Державина, Марины Цветаевой и Никиты Заболоцкого, художников Ивана Шишкина и Николая Фешина, музыкантов Олега Лундстрема и Михаила Плетнёва. Классик татарской поэзии Габдулла Тукай, поэт-герой Муса Джалиль, композиторы Фарид Яруллин, Салих Сайдашев, Назиб Жиганов, София Губайдулина составили славу татарской культуры.
В республике значительное количество библиотек, клубов, музеев театрально-зрелищных учреждений и кинотеатров, а также памятников архитектуры, истории и культуры.
Казанский Кремль является объектом Всемирного наследия ЮНЕСКО как «выдающийся образец синтеза татарского и русского влияния в архитектуре», а в 2014 году список Всемирного наследия ЮНЕСКО пополнил древний город Болгар, где предки татар в 922 году приняли ислам. Восстановление древнего Болгара – часть большого Комплексного проекта «Культурное наследие Татарстана: остров-град Свияжск и Древний Болгар».
Содействие этнокультурному развитию всех народов, проживающих в Татарстане, – одно из ключевых направлений государственной политики в сфере культуры.
Межнациональное и межконфессиональное согласие является одним из трех важнейших составляющих успешности Татарстана как субъекта Российской Федерации, наряду с высоким уровнем экономического развития и общественно-политической стабильностью.
Создание положительного имиджа Татарстана, пропаганда культуры Татарстана, сохранение татарского языка и культуры, поддержка соотечественников – еще одно ключевое направление деятельности в сфере культуры.
Широкую известность получили международные фестивали: оперный им. Ф.И. Шаляпина, классического балета им. Р.Х. Нуриева, современной музыки «Европа – Азия», театральный фестиваль тюркских народов «Науруз», открытый республиканский телевизионный молодежный фестиваль эстрадного искусства «Созвездие-Йолдызлык».
Особой гордостью Татарстана является Казанский международный фестиваль мусульманского кино, который стал площадкой для конструктивного диалога между представителями различных культур и конфессий.
Значимость вклада татарского народа в современную мировую культуру подтвердило и решение Международной организации тюркской культуры ТЮРКСОЙ о статусе Казани как культурной столицы тюркского мира.
Основное туристическое направление Татарстана – это экскурсии по историческим местам республики. Множество архитектурных комплексов, храмов находятся в прекрасном состоянии и всегда открыты для гостей.
Остров-град Свияжск (прототип пушкинского острова Буяна), тысячелетний город Елабуга и Великий Болгар, очарование природы Приволжья – всё это – культурное наследие республики: изящное переплетение татарских и русских традиций, православия и ислама.
Много на Волге достопримечательностей и другого рода. Это и Романовский железнодорожный мост, построенный в начале прошлого века, и любимый учеными всего мира Печищинский геологический разрез, каждый слой которого соответствует отложениям разных эпох, и Волжские (Сюкеевские) карстовые пещеры в районе Камского Устья, и старинные прибрежные храмы со своими удивительными историями.
Площади, Республика Татарстан, Россия — Туристер.Ру
16 туристов было здесь
Сегодня площадь Тысячелетия — это самая крупная и известная, а также одна из двух главных площадей Казани (наряду с площадью Свободы)….
35 туристов было здесь
Одним из самых интересных, красивых и наиболее часто посещаемых туристами объектов в г. Казани является Дворцовая площадь, также называемая иногда…
6 туристов было здесь
Площадь Свободы — самая большая площадь Казани, одна из двух главных площадей столицы Татарстана (вместе с площадью Тысячелетия). Площадь…
34 туристa были здесь
Площадь Тукая — сердце Казани. Названа она так в честь татарского публициста, поэта и общественного деятеля Габдуллы Тукая….
Площадь Султан-Галиева расположилась в центральной части Казани. Это современное и комфортабельное место, куда любят приходить горожане,…
Николаевская площадь располагается в Вахитовском районе Казани, в исторической части города. Ранее носила название Кузнечная. В …
5 туристов было здесь
Площадь 1 мая изначально называлась Спасская, затем Ивановская и Александровская. Это древнейшая площадь города. Располагается она…
Площадь Соцгорода находится в Авиастроительном районе Казани. Жители чаще называют ее «Площадь у ДК Ленина» или…
1 турист был здесь
Площадь Вахитова находится в центре Казани на границе двух районов Вахитовского и Приволжского. Имеет интересное расположение,…
Площадь Моторостроителей находится в Авиастроительном районе, в поселке-микрорайоне Караево. Располагается у пересечения улиц…
татарский | люди | Britannica
Татар , также пишется Татар , любой представитель нескольких тюркоязычных народов, которые в совокупности насчитывали более 5 миллионов в конце 20 века и жили в основном на западе центральной России вдоль центрального течения реки Волги и его приток Кама, а оттуда на восток до Уральских гор. Татары также проживают в Казахстане и, в меньшей степени, в Западной Сибири.
Название татар впервые появилось у кочевых племен, живших на северо-востоке Монголии и в районе озера Байкал с V века нашей эры.В отличие от монголов, эти народы говорили на тюркском языке и могли быть родственниками половцев или кипчаков. После того, как различные группы этих тюркских кочевников вошли в состав армий монгольского завоевателя Чингисхана в начале 13 века, произошло слияние монгольских и тюркских элементов, и монгольские захватчики России и Венгрии стали известны европейцам как татары (или Татары).
Подробнее по этой теме
Россия: татарское правление
После непродолжительной попытки возродить древние центры Булгара и Крыма, Джусиды (семья Джочи, сына Чингисхана, унаследовавшего…
После распада империи Чингисхана татары стали особенно отождествлять себя с западной частью монгольского владения, которая включала большую часть Европейской России и называлась Золотой Ордой. Эти татары были обращены в ислам суннитов в 14 веке. Из-за внутренних разделений и различного внешнего давления Золотая Орда распалась в конце XIV века на независимые татарские ханства Казань и Астрахань на Волге, Сибирь в Западной Сибири и Крым.Россия завоевала первые три из этих ханств в XVI веке, но Крымское ханство стало вассальным государством турок-османов, пока не было присоединено к России Екатериной Великой в 1783 году.
В своих ханствах у татар сложилась сложная социальная организация. , и их дворянство сохранило свое гражданское и военное руководство до русских времен; отдельные классы простолюдинов были купцами и земледельцами. Во главе правительства стоял хан передового татарского государства (Казанского ханства), часть семьи которого в XVI веке по прямому сговору присоединилась к русской знати.Это расслоение в татарском обществе продолжалось до русской революции 1917 года.
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишитесь сейчасВ 9-15 веках татарская экономика основывалась на смешанном земледелии и скотоводстве, которое продолжается до сих пор. У татар также сложились традиции изготовления изделий из дерева, керамики, кожи, ткани и металла, и они давно известны как торговцы. В течение 18 и 19 веков они заработали привилегированное положение в расширяющейся Российской империи в качестве торговых и политических агентов, учителей и администраторов недавно завоеванных территорий Центральной Азии.
Более 1,5 миллиона казанских татар по-прежнему проживают в Приволжском и Уральском регионах, и они составляют около половины населения Республики Татарстан. Теперь они известны как волжские татары и являются самой богатой и наиболее развитой в промышленном отношении группой татар. Еще почти миллион татар проживает в Казахстане и Средней Азии, в то время как сибирские татары, которых насчитывается всего около 100 000 человек, живут разбросанными по Западной Сибири.
Крымские татары в Новое время имели свою историю.Они легли в основу Крымской Автономной Советской Социалистической Республики, созданной Советским правительством в 1921 году. Эта республика была распущена в 1945 году, однако после того, как советский лидер Иосиф Сталин обвинил примерно 200000 крымских татар в сотрудничестве с немцами во время войны. Вторая Мировая Война. В результате крымские татары были массово депортированы в Узбекистан и Казахстан, где использование татарского языка было запрещено. Они восстановили свои гражданские права в 1956 году в рамках программы десталинизации Никиты Хрущева, но им не разрешили вернуться в Крым, который был включен в состав украинского С.S.R. в 1954 году. Лишь в начале 1990-х многие крымские татары, воспользовавшись распадом советской центральной власти, начали возвращаться в Крым после почти пяти десятилетий внутреннего изгнания. В начале 21 века их насчитывалось около 250 000 человек.
Тартария
«Тартария» перенаправляется сюда. Чтобы узнать о румынской деревне Тэртэрия, см. Сэлиштя. Чтобы узнать о субъекте Российской Федерации, ранее известном как Татария, см. Татарстан.Тартария или Великая Тартария (латинское: Tataria или Tataria Magna ) — название, которое европейцы использовали со времен средневековья до двадцатого века для обозначения Великой степи, то есть большого участка северной и центральной частей страны. Азия, простирающаяся от Каспийского моря и Уральских гор до Тихого океана, населенная в основном тюрками, монголами, а также некоторыми казаками русского происхождения, гражданами Монгольской империи, которых в целом называли «татарами», т. Е.е. Татары. В него вошли нынешние районы Сибири, Туркестана, Монголии и Маньчжурии.
География и история
Тартария часто делилась на части с приставками, обозначающими имя правящей державы или географическое положение. Таким образом, Западная Сибирь была Москвича или Русской Тартарии , Синьцзяна и Монголии были Китайской или Катайской Татарии , Западный Туркестан (позже Русский Туркестан) был известен как Независимая Тартария , а Маньчжурия была Восточная Тартария .
По мере того, как Российская империя расширялась на восток, и европейцы стали узнавать все больше о Татарии, термин вышел из употребления.
европейских областей к северу от Черного моря, населенных тюркскими народами, были известны как Малая Тартария.
«Комулская пустыня в Татарии» упоминается Иммануилом Кантом в его «Наблюдениях за чувством прекрасного и возвышенного», как «великое далеко идущее одиночество».
Восточная Тартария
Восточная Тартария и Приморская Тартария — старые названия населенных маньчжурцами территорий, простирающихся от места слияния реки Амур с рекой Уссури до острова Сахалин.Сейчас это Приморский край с Владивостоком в качестве регионального административного центра.
Эти земли когда-то были заняты племенами мохэ и чжурчжэнями; а также различными старыми королевствами, включая Годжосон, Балхэ, Ляо и (киданьские) королевства.
Согласно летописи династии Мин Шэн-Ву-Чи («Наша династия известна военными осознаниями»), на этой земле основались племена тунгус вэджи, варка и курка. Позже они были объединены в Маньчжурскую империю Цин с Нурхачи в качестве их лидера и основателя.Эти земли были потеряны для России по Пекинскому договору.
Ближайший к этой земле находится остров Ку-Е-Дао (китайский: 庫 頁 島; пиньинь: Kùyèdǎo ) или остров Фу-Санг (хангыль: 후상), более известный как Карафуто или Сахалин; Не так давно русские археологи нашли здесь остатки древних городов со стенами и замками. Они могут соответствовать древней маньчжурской нации, или, возможно, во времена монголов или тунгусов, или царству Балхае.
Эти земли посещали японские исследователи Мамия Ринзо и другие, которые рассказывали о различных важных городах и портах, таких как Хайшенвэй (современный Владивосток).С этих земель и близлежащего Хулуна (Амурская область) японцы заявили свои права на предков из Северной Азии, которые заселили Северную Японию.
Другими древними городами в регионе являются: Тетюхе (ныне Дальнегорск) и, вероятно, Деленг, важный торговый имперский пост, согласно некоторым источникам.
Татария в художественной литературе
В романе Владимира Набокова Ада , Тартария — название большой страны на вымышленной планете Антитерра. Россия является приблизительным географическим аналогом Татарии на Терре, двойном мире Антитерры, очевидно идентичном «нашей» Земле, но вдвойне вымышленном в контексте романа.
В последней опере Пуччини «Турандот» отец Калафа Тимур — свергнутый король татар.
В романах Филиппа Пуллмана «Темные материалы» главные герои Европы часто выражают страх перед татарами — термин, очевидно, относящийся ко многим азиатским расам, поскольку действие повествования происходит далеко от Монголии.
В «Макбете» Уильяма Шекспира ведьмы включают губы татар в свое зелье.
В фильме Мэри Шелли « Франкенштейн » доктор Франкенштейн преследует монстра «среди дебрей Татарии и России», хотя он все еще уклонялся от меня, я когда-либо следил за ним.«
В книге Чарльза Диккенса «Большие надежды» Герберт Покет описывает Эстеллу Хэвишем как татарину, потому что она была «жесткой, надменной и капризной до последней степени и была воспитана мисс Хэвишем, чтобы отомстить всему мужскому полу». [1]
В «Крысолове Гамлена» Роберта Браунинга Крысолов упоминает Тартарию как одно из своих свидетельств мэру Хамельна в борьбе с вредителями. «В Тартарии в июне прошлого года я освободил Чама от его огромных скоплений мошек»;
В своей небольшой работе с Э.Хоффманн Прайс, «Через врата Серебряного ключа», Г.П. Лавкрафт кратко упоминает Тартарию: «На их замаскированных головах теперь, казалось, покоились высокие митры неопределенного цвета, что странно напоминало те, что были на безымянных фигурах, вырезанных забытым скульптором вдоль холма живые скалы высокой запретной горы в Тартарии … »
«Рассказ сквайра» из сериала « Кентерберийские рассказы» Джеффри Чосера «» расположен в королевском дворе Тартарии.
В «Путешествиях Гулливера» Джонатана Свифта эпонимический герой дважды упоминает о своих путешествиях в Тартарию и предполагает, что тогдашние современные географы Европы «совершили большую ошибку, предполагая, что между Японией и Калифорнией не было ничего, кроме моря. мое мнение, что должен быть баланс земли, чтобы уравновесить большой континент Тартария… «
См. Также
Ссылки и дополнительная литература
Цитаты
Питер Флеминг: Компания одного (1936) и Новости из Тартарии (1936), позже опубликованные вместе как «Путешествие в Тартарию».
Внешние ссылки
Тартария: Империя, скрытая историей или открытая невежеством?
Во время недавних блужданий по Интернету я наткнулся на очень специфический сабреддит: «Тартария». Поначалу я думал, что этот субреддит посвящен какому-то фантастическому или вымышленному миру в стиле стимпанк, но после нескольких постов я понял, что на самом деле все дело в теории заговора относительно древнего королевства, игнорируемой или намеренно скрытой историками.Большинство постов там показывают здания в европейском стиле в Соединенных Штатах, построенные, предположительно, с использованием «татарских» технологий, или спекулируют на художественных репрезентациях «утраченных технологий».
Мне как историку эти теории казались бессмысленными и абсурдными. Однако я не мог перестать гадать, о чем все это и откуда могла взяться теория. После некоторого исследования я обнаружил, что это симптом серьезной проблемы: дезинформация, недоверие к науке и ее методам, а также вера в лженауку.Кое-где в теории в целом встречаются и другие проблемы, и я скоро расскажу о них, но обо всем по порядку.
Происхождение: мое невежество так же достойно, как и ваше знание
Я сделаю оговорку: когда я говорю «незнание», я просто имею в виду отсутствие знаний о предмете, а не ad-hominem по отношению к людям, верящим в эти теории заговора. Что ж: я проследил большую часть обсуждения Тартарии до блога под названием «Украденная история» (который с тех пор был удален).
Предпосылка блога неплохая, он утверждает, что историю пишут победители, поэтому он заставляет людей находить различные точки зрения и факты: то, что поощряют историки, ища как можно больше источников. Тем не менее, поскольку это открытый форум, почти все предположения обсуждаются серьезно, без особых доказательств. Вот когда дела идут плохо.
По сути, некоторые люди заметили, что до 18-19 веков карты включали регион под названием «Тартария» или «Великая Тартария» на востоке России, в Средней Азии и Сибири.Они объединяют это с цитатами из Британской энциклопедии (работа 18 века) в статьях вроде , эта , чтобы утверждать, что в истории существует скрытая Империя.
Это свидетельствует о полном отсутствии исторического контекста. Европейские географы не имели глубоких знаний об этом регионе и его народах — кочевой природе тюркского и монгольского населения, бродившего по степям. Например, на картах просто обозначен регион, где раньше бродили эти кочевые народы, а не королевство по образцу западного.По мере того как географы узнавали больше об этом регионе, термин «татарин / татарин» утратил свое значение в качестве обобщающего термина для кочевого населения региона. Другие элементы, такие как «татарский язык», флаг и герб, которые были обобщены от одного племени ко всем другим, живущим в этом регионе, теоретики заговора считают доказательством существования такой могущественной Империи.
Становится хуже. Еще одним предполагаемым доказательством того, что академическая история является мистификацией, является несоответствие между изображениями людей, таких как Чингисхан, Бату-хан и Тамерлан, в 15-18 веках и более современными (примеры, которые они используют, на самом деле, такие же старые, только они картины, сделанные придворным живописцем настоящего правителя).Только «старые» изображения, которые считаются более точными, являются белоснежными версиями монархов: поскольку европейские художники, создавшие их, не знали этническую принадлежность, которую они представляли, поэтому они просто представляли Чингисхана как обычного западного короля. Это не просто незнание, но и сдержанный расизм.
Азиатские и европейские изображения правителей Средней Азии / Фотомонтаж изображений с сайта Stolenhistory.org
Но на этом не заканчивается. Помните, что я сказал о зданиях в Соединенных Штатах? Что ж, основываясь на цветовой кодировке на двух старых картах и плохо интерпретированной латыни, есть мнение, что Тартария имела владение в Северной Америке .Вот так. Связывая все это вместе, теоретики заговора полагают, что большинство архитектурных стилей и технологий, связанных с Западной Европой, в том числе в Европе и за ее пределами, на самом деле являются татарскими. Старые рисунки и картины, изображающие, какими будут технологии в будущем, стиль Жюля Верна, изображающие людей в летных костюмах и причудливых вертолетах в стиле стимпанк, приписываются Тартарии, подразумевая, что Империя обладала передовыми технологиями, которые были просто скрыты, уничтожены или изменены европейскими завоевателями. .
Есть целые дискуссии о , как старые карты изменяли свою форму странным образом (полностью игнорируя, насколько сложной и неточной была картография без современного оборудования), приписывая такие изменения сильным наводнениям и тому подобному. Я мог бы продолжать и продолжать, но примеров, которые я здесь привел, достаточно, чтобы показать, как работает эта «скрытая история» — путем выбора вишен и распознавания искаженных образов.
1652 Карта мира / Stolenhistory.org
Заключение: все дело в контексте …
Как бы снисходительно я ни звучал, излагая эти теории, я не думаю, что люди на подобных форумах должны просто молчать.Опрос — фундаментальная часть науки; все великие открытия происходят из желания понимать больше, осознания того, что вещи, которые мы знаем, неполны. Любопытство и желание улучшить наше понимание малоизученных тем — это хорошо, но это не должно быть просто бездумным предположением. Вот почему существует научный метод: вы перекрестно проверяете факты и информацию, рецензируете статьи, обсуждаете теории с экспертами на конгрессах и семинарах, проводите дискуссии в академических кругах. Действительно, интересно видеть, как люди стремятся к знаниям ради самих себя.
Чего мне не хватает, так это понимания того, как создаются знания. . Иногда мы думаем, что то, что мы знаем, считается само собой разумеющимся — мы слушаем наших учителей в школе и читаем наши книги, уже думая, что это всегда было там, и что все согласны с тем, что с тех пор считается фактами. Вера в заговоры и псевдонауку часто возникает с того момента, когда люди осознают, что знания могут быть оспорены — и, кстати, оспариваются постоянно! Что вы имеете в виду, есть альтернативные объяснения тому, чему меня учили? Что еще может быть ложью? Вот и они, отправляясь искать знания, но без метода.
Метод важен, или, скорее, фундаментален, и, возможно, нам нужно больше узнать об этом, чем узнавать о том, что было открыто на данный момент. Каждый может заниматься наукой и производить знания, да, абсолютно, но без метода мы в конечном итоге соединяем точки, совершенно не связанные друг с другом.
Тартария показывает, как простое незнание культуры и истории других людей может привести к огромным недоразумениям. В Бразилии, где я вырос, азиатская история и русская революция — темы, которые почти не затрагиваются, и только потому, что они относятся к большему европейскому сценарию; Европейской истории уделяется наибольшее внимание в западном образовании, при этом мало внимания уделяется истории Африки и Азии.Без учебы я мог бы так же легко попасться на это.
Если вы думаете, что такая теория безвредна, просто примите во внимание неудачи, которые мы переживаем, потому что есть сторонники плоской земли и люди, которые считают, что нацисты были левым крылом. Подобные теории заговора замедляют развитие науки, распространяют дезинформацию и поддерживают бредовое мышление. Во всяком случае, это отвлекает ученых от продолжения исследований по расширению наших текущих знаний, заставляя их доказывать то, что уже широко известно.
Как и большинство сегодняшних проблем, ключом к решению этой общей дискредитации науки и широко распространенного теоретизирования заговора может быть только один: образование, хорошее, образование. Царство Тартария и все его технологии могут быть ненастоящими, но теории заговора, которые наносят ущерб науке и доверию к академической истории, вполне реальны.
Рекомендуемое изображение: Карта Тартарии 1754 г. / Stolenhistory.org
растений | Бесплатный полнотекстовый | Гречневая крупа в питании человека
1.Введение
Гречиха татарская (Fagopyrum tataricum Gaertn.) Возникла в западном Китае [1] и выращивается в горных районах Китая, Бутана, северной Индии и Непала [2,3]. В тех же странах в регионах с менее суровыми климатическими условиями гречиха по-татарски выращивается вместе с гречкой обыкновенной (рис. 1). В Европе татарская гречиха традиционно культивируется и широко используется в Люксембурге и в прилегающих районах Бельгии и Германии, а также известна в Словении и Италии [3,4,5].Гречиха по-татарски также традиционно выращивается в Боснии и Герцеговине, а также является смешанной с гречкой обыкновенной. Недавно сообщалось, что в Вермланде, Швеция, ежегодно выращивается более 100 гектаров татарской гречихи [6]. Сбор образцов татарской гречихи на словенских полях начался в конце 1970-х годов. В то время его культивировали всего несколько фермеров, и вместо этого на многих гречихных полях он сосуществовал с обычной гречкой, но как сорняк. Многолетний исследовательский проект Министерства сельского хозяйства Люксембурга стимулировал исследования, выращивание и развитие татарской гречихи в Европе [3,4].Гречка обыкновенная и татарская имеют разные ростовые характеристики [7]. Известно, что гречиха винная устойчива к воздействию холода из-за ее эпигенетической регуляции метилированием ДНК [8]. Кроме того, она более засухоустойчива, чем гречка обыкновенная. Действительно, гречиха татарская обладает признаками засухоустойчивости, тогда как гречиха обыкновенная обладает способностью избегать засухи [9]. Род Fagopyrum включает 21 вид [1,10]. Два из них, Fagopyrum esculentum и F. tataricum, используются в питании человека, а дикие виды Fagopyrum cymosum используются в традиционных китайских лекарствах для человека и в ветеринарии.Дикие сородичи культурной гречихи распространились по горным районам юго-западного Китая [11], где недавно был обнаружен новый самосовместимый вид, описанный М. Чжоу и Ю. Тангом как Fagopyrum longistylum [12]. F. longistylum и другие виды дикой гречихи могут служить донорами генов при селекции культурных видов гречихи [13]. Татарская гречиха включает генотип, называемый «рисово-татарская гречка». Удаление шелухи (шелухи) этого варианта легче по сравнению с другими вариантами татарской гречки, поскольку шелуха более тонкая и хрупкая [14].Сравнительный анализ транскриптомов этих двух генотипов татарской гречихи показал, что между ними по-разному экспрессируются 9250 генов. К ним относятся различия в регуляторных и структурных генах, которые влияют на химические компоненты клеточной стенки. Поскольку у татарской гречки обычно более толстая и прочная шелуха, чем у обычной гречки, этот вариант рисово-татарской гречки важен для облегчения лущения и получения крупы. Образцы винной гречихи обнаруживают множественные участки одомашнивания, как было показано последовательным секвенированием большого количества татарской гречки. образцы [12].Внутривидовые скрещивания с участием гречихи татарской привели к появлению нескольких новых гибридных видов, среди которых, вероятно, наиболее известен Fagopyrum giganteum Krotov, который первоначально был определен Кротовым и Драненко на Устимовской опытной станции в Украине [15,16]. Дикий вид Fagopyrum homotropicum был важным источником для развития самоопыления у культивируемых видов гречихи [17]. Потребление гречихи оказывает защитное действие против ряда хронических заболеваний, включая гипертонию, ожирение, сердечно-сосудистые заболевания и образование камней в желчном пузыре [18].Эти эффекты в основном объясняются устойчивым крахмалом, белком и фенольными веществами в зерне гречихи.2. Устойчивый крахмал
Содержание амилозы в крахмале является основой появления ретроградного крахмала [19]. Амилоза является исходным материалом для получения устойчивого крахмала с помощью различных гидротермальных обработок [20]. Резистентный крахмал — это часть крахмала, которая не переваривается человеческими ферментами до попадания в толстую кишку. На количество резистентного крахмала влияет состав крахмала с точки зрения высокого содержания в нем амилозы и в зависимости от экологических и генетических факторов [21,22,23].У сорта гречихи Татарский Xinong9920 пиковая вязкость крахмала составляла 2121 сП, а в тех же условиях пиковая вязкость сорта Xinong9940 была значительно меньше, а именно 1928 сП [22]. В опыте с фосфорными удобрениями это было у татарской гречихи при дозе Р 75 кг / га, в крахмале кажущаяся амилоза 24,7% от общего количества крахмала (27,0% в неудобренных), а в другом эксперименте 27,4% (28,6% в неудобренных) в общем количестве крахмала [23]. Удобрение татарской гречки фосфатом влияет на ее рост, развитие и качество.Zhang et al. [23] показали, что внесение удобрений с различным содержанием фосфора (P) (от 15 до 135 кг / га) влияет на характеристики крахмала из татарской гречихи. Увеличение содержания Р сначала уменьшалось, а затем увеличивалось кажущееся содержание амилозы и средний диаметр гранул крахмала. Эти эффекты повлияли на ретроградацию крахмала, процесс, посредством которого после смешивания крахмала с горячей водой дезагрегированные цепи амилозы и амилопектина затем подвергаются рекомбинации при охлаждении с образованием более упорядоченной структуры.Скорость ретроградации татарской гречневой крахмальной пасты увеличивалась в течение первых 8 ч после начала охлаждения, а затем постепенно стабилизировалась. Повышенное содержание амилозы способствует быстрой ретроградации, а медленная ретроградация крахмала — это эффект молекул амилопектина. Соответственно, крахмал из татарской гречихи, подвергнутый обработке с высоким содержанием фосфора, имеет большое количество разветвленных длинных цепей амилопектина, поскольку высокая концентрация длинных цепей in situ происходит в процессе реформирования спиралей крахмала.Гидротермальная обработка крахмала зерна гречихи по-винному увеличивает уровень медленно усваиваемого крахмала за счет ретроградного крахмала [24]. Зерна гречихи имеют относительно небольшие гранулы крахмала, причем содержание амилозы в крахмале зерна выше, чем в зерновых [25]. Поскольку татарская гречка имеет высокое содержание флавоноидов, которые могут взаимодействовать с молекулами крахмала, татарская гречка может использоваться для производства продуктов с низким гликемическим индексом [22]. Повышенный резистентный крахмал получают путем варки гречневой крупы [25], а также обработкой холодной плазмой и комплексообразованием кверцетина [26].Это переваривание резистентного крахмала является частью наших пищевых волокон и действует как пребиотик. Различные сорта татарской гречки имеют разные характеристики крахмала, и поэтому необходимо учитывать это при переработке гречихи [22]. Условия выращивания гречихи, наличие органических веществ во время наполнения и созревания гречневого зерна, а также наследственные характеристики гречихи — все это имеет важное влияние на размер гранул крахмала и содержание амилозы крахмала.Образцы гречихи, подвергнутой гидротермической обработке, содержали до 4% ретроградного крахмала по сравнению с необработанной и нагретой сухим способом гречихи, которая содержит только около 1% устойчивого крахмала в виде сухого вещества [19,27,28]. Прогресс в исследованиях обычного и винного крахмала. размер и форма зерен рассмотрены в [22]. Более низкий гликемический индекс крахмала и инсулиновый индекс после нагревания связывают с образованием крахмала, устойчивого к амилазе [29]. Культуры молочнокислых бактерий и бифидобактерий могут использоваться для приготовления напитков из обычных продуктов, богатых крахмалом гречихи.В Словении из ферментированного теста также готовят изделия из татарского гречневого хлеба [30,31].3. Белок
Зерно гречихи считается псевдозлаком с высокой питательной ценностью из-за его белкового состава. Хотя зерно гречихи имеет низкое содержание белка (гречиха обыкновенная, 10,6 г / 100 г сухой массы; гречиха татарская, 10,3 г / 100 г сухой массы), она имеет сбалансированный аминокислотный состав с высоким содержанием незаменимых аминокислот, таких как как лейцин и лизин (гречка обыкновенная: 6,92, 5.84 г / 100 г белка; Гречка по-винному: 7,11, 6,18 г / 100 г белка; соответственно) [2]. Высокое содержание белка, флавоноидов и микроэлементов в некоторых фракциях помола гречихи предполагает их использование в специальных диетических продуктах [32]. Белок зерна гречихи также может содержать Se [33], который является важным микроэлементом в питании человека. Были изучены различные гидротермальные обработки зерна гречихи, чтобы определить влияние уровней полифенолов на усвояемость белка [29]. В модельной системе на крысах во время гидротермальных обработок наблюдались значительные взаимодействия между полифенолами и белком.Эти взаимодействия снижают переваривание белка зерна гречихи в тонком и толстом кишечнике. Однако микробные процессы в толстой кишке улучшили усвояемость протеина гидротермально обработанной гречихи, которая в противном случае блокировалась полифенолами [29]. Авторы установили, что природные полифенолы, содержащиеся в шелухе гречихи, снижают истинную усвояемость белка зерна гречихи, но не влияют отрицательно на биологическую ценность. Как сообщает Ikeda et al.[34], дубильная кислота и катехин оказывают значительное ингибирующее действие на in vitro пептическое и панкреатическое переваривание гречишного глобулина. Икеда и др. [34], Икеда и Кишида [35] изучали усвояемость белка зерна гречихи in vitro и влияние вторичных метаболитов гречихи. Литературные данные указывают на то, что белок зерна гречихи может снижать уровень холестерина в сыворотке за счет увеличения фекальной экскреции стероидов, которая индуцируется связыванием стероидов с непереваренным белком.Согласно Ма и Сионгу [36], устойчивые к пищеварению пептиды в значительной степени ответственны за выведение желчных кислот. Эти эффекты, скорее всего, связаны с ограниченной усвояемостью белка зерна гречихи. Поскольку зерно гречихи не содержит белков глютена, оно используется для приготовления пищи пациентам с глютеновой болезнью [37,38]. Хотя аллергия на гречку не очень распространена, сообщалось об аллергических расстройствах, связанных с употреблением продуктов на ее основе [39,40,41]. Низкомолекулярные белки зерна гречихи, связанные с такой аллергией, находятся в зародышах зерна, а не в эндосперме [42].При традиционной гидротермальной подготовке гречневой крупы происходит миграция веществ из околоплодника зерна в крупу [43,44]. При переработке зерна гречихи для производства различных пищевых продуктов возможны различные взаимодействия между составляющими, особенно во время гидротермальной обработки. Jin et al. [45] предположили, что зерно гречихи можно обрабатывать для улучшения перевариваемости белка и повышения биологической активности белка гречихи обыкновенной и татарской. Таким образом, гречишное зерно можно использовать в качестве источника белка на растительной основе для улучшения глобального снабжения белком.4. Минеральные элементы
Зерно и зерновые продукты гречихи по-винному имеют высокий уровень минеральных элементов [3]. Эти уровни зависят от процесса помола, при этом самые высокие уровни минеральных элементов содержатся в отрубях; меньше в темной муке и меньше всего в тонкой, легкой муке. Уровни Rb и Ag в обычной гречке выше, чем в татарской гречке, хотя уровни других минеральных элементов (например, Se, Zn, Fe, Co, Ni, Sb, Cr, Sn) выше в татарской гречке. Однако уровни всех изученных элементов в муке из листьев гречихи по-татарски намного выше, чем в зерне или измельченных зерновых продуктах.Настои из листьев гречихи по-винному содержат снижающиеся уровни (в порядке убывания) Zn, Cu, Cr, Ni, Pb и Cd [46]. В этом исследовании сообщалось, что концентрации Cr в настоях из цельного растения, а также из зерновых отрубей и зародышей находятся в диапазоне от 2,5 мг / кг до 3,2 мг / кг. В образцах, собранных на коммерческих рынках Китая, видно, что используемая часть завода и методы обработки влияют на концентрации этих металлов в продуктах. Для содержания Cr гречиху по-татарски для этих настоев необходимо выращивать в чистой среде.Использование зеленых частей татарской гречки или отрубей в настоях показало, что по сравнению с напитками из круп, эти настои содержат более высокие концентрации Cd (0,5–1,2 мг / кг), Pb (0,3–0,4 мг / кг), Cu (5– 8 мг / кг) и Zn (30–50 мг / кг) [46]. Если листья растений и шелуха зерна загрязнены частицами почвы, возможно, загрязненная почва является прямым источником загрязнения татарской гречихи металлами. С другой стороны, ионы металлов могут связываться с флавоноидами, что может способствовать абсорбции металлов растениями татарской гречихи и, таким образом, их распределению в частях, богатых флавоноидами.Согласно Li et al. [46], уровни Pb в листьях не связаны с содержанием флавоноидов или общим содержанием фенолов. Татарская гречневая мука из Люксембурга, в Европе, содержала только 0,32 мг / кг Cr в темной муке и только 0,10 мг / кг Cr в муке тонкого помола [3]. Обычная гречиха может накапливать Al в своих листьях, хотя это хранилище Al не выражается в зерне. Это, по-видимому, связано с тем, что нет транспорта Al через флоэму, и Al не является подвижным после его накопления в листьях.Накопленный Al в старых листьях, по-видимому, происходит из корней, и поэтому зеленые части растений гречихи можно использовать для удаления Al из почвы [47,48,49]. может накапливать значительное количество Se. Таким образом, он может быть источником селена в рационе человека [33,50]. Салициловая кислота может повышать уровень селена в тканях растений [51]. Следует отметить, что пациенты с тяжелой инфекцией COVID-19 имеют дефицит витаминов D и Se. Действительно, поскольку Se, по-видимому, усиливает цитотоксические эффекторные клетки, дефицит Se является возможным фактором риска смертности от COVID-19 [52].5. Фенольные вещества
Yu et al. [53] сравнили содержание рутина и кверцетина в 44 образцах зерна и проростков татарской гречихи из Китая, Непала, Бутана, Индии, Японии, Пакистана и Словении. Они были очень разными в зависимости от происхождения (таблица 1). Образцы из Непала имели самые высокие концентрации рутина в зерне (13,3 г / кг) и проростках (54,4 г / кг). Для кверцетина ростки содержали в 10–90 раз больше, чем зерна [53]. Таким образом, проростки татарской гречки обладают большим потенциалом для производства флавоноидов и для получения функциональных продуктов, богатых флавоноидами.По данным Клыкова и соавт. [54], в пробах с Дальнего Востока России зерно гречихи обыкновенной содержало 0,1% рутина по весу, гречихи татарской 2,4% и гречихи cymosum 1,1%. Во время полного цветения надземные части гречихи обыкновенной содержали от 3,1% до 3,8% рутина, давая от 92 до 121 кг рутина / га; то же самое для татарской гречихи — от 4,1% до 4,4% рутина, для 107–129 кг рутина / га, а для гречихи cymosum — ~ 4,1% рутина на ~ 83 кг рутина / га [54]. Следует отметить, что татарская гречиха с темным покровом зерна содержит больше рутина, чем татарская гречиха с другим цветом зерна [55].Кроме того, уровень озона в атмосфере может повлиять на биосинтез фенольных веществ в татарской гречке [56,57,58]. Помол зерна татарской гречихи и смешивание муки с водой приводит к образованию кверцетина как продукта разложения рутин под действием рутинозидазы (рис. 2) [59,60,61,62,63]. Однако перегретый пар или насыщенный пар можно использовать для инактивации ферментов, разлагающих рутин, в гречневой муке менее чем за 90 секунд. Напротив, при сушке в дальней инфракрасной области эти ферменты, разрушающие рутин, сохраняются при 150 ℃ в течение 40 минут [64].Попавший внутрь кверцетин может преодолевать гематоэнцефалический барьер и накапливаться в тканях мозга [65]. Действительно, была установлена важная биоактивность кверцетина и его производных не только в кровеносных сосудах, мышцах и желудочно-кишечном тракте, но и в головном мозге. Кверцетин и другие фенольные соединения были выделены из образцов стула людей, которые ели пищу, богатую фенольными веществами [65]. Присутствие фенольных веществ в толстой кишке может снизить вирусную нагрузку в стуле. В татарской гречке образование комплекса кверцетина с молекулами крахмала влияет на усвояемость крахмала in vitro и появление резистентного крахмала, изменяя таким образом физико-химические свойства Гречишный крахмал татарский [66].Эффекты этого комплексообразования кверцетин-полифенол указывают на то, что пищевые продукты на основе татарской гречихи будут демонстрировать более низкую усвояемость. Действительно, кверцетин в татарской гречке может снизить массу тела, уровень триацилглицеринов в сыворотке и липопротеинов низкой плотности. Было показано, что у крыс диета с 0,1% кверцетина оказывает значительное влияние на снижение концентраций липопротеинов низкой плотности в сыворотке без такого воздействия на липопротеины высокой плотности. Также было показано, что винная гречка предотвращает увеличение массы тела и отложение жира при потреблении большого количества жиров у крыс, хотя, с другой стороны, сообщалось, что это защищает от стеноза печени [67].Гречневая диета также может снизить уровень инсулина и уменьшить непереносимость глюкозы у людей [19]. Эксперименты на крысах с гречневой крупой также подтвердили сложность воздействия микробиоты кишечника. Действительно, Peng et al. [67] предположили, что связь между прибавкой в весе и микробиотой кишечника очень сложна, и здесь необходимы дальнейшие исследования. Интересно, что было показано, что обогащенные рутином экстракты татарской гречневой муки обеспечивают лучшее пероральное всасывание флавоноидов с фенольными веществами. в крови обнаруживается дольше, чем при использовании стандартного рутина, и даже дольше, чем для экстракта муки из нативной татарской гречихи [45].Рутин в большей степени связан с другими веществами и структурами зерна. Действительно, экстракция рутина из необработанной муки из зерна татарской гречихи показала 0,57 г рутина / 100 г муки, в то время как автоклавирование дало 3,03 г / 100 г муки, кипячение дало 2,97 г рутина / 100 г муки, а пропаривание дало 2,50 г / 100 г муки. г муки [45,68]. Дза и др. [68] также изучали условия твердо-жидкостной экстракции татарской гречихи, где они указали, что экстракция фенольных соединений из татарской гречневой муки может быть проведена на следующем этапе. Рутин и кверцетин в зерне татарской гречихи оказывают влияние на физико-химические свойства крахмала. приготовление еды.Энтальпия старения ретроградного крахмала снижается, а вязкость татарского гречневого крахмала и пасты увеличивается. Связывание крахмал-фенольные соединения сильнее, чем у комплекса крахмала и йода. Крахмал желатинизирован и ретрограден, а на морфологию влияют кверцетин и рутин [69]. Среди фенольных соединений некоторое количество ресвератрола было зарегистрировано в муке из зерна гречихи обыкновенной [70]. Немцова и др. [71] сообщили о транс-ресвератроле от 1,0 мг / кг до 1,7 мг / кг для зерна гречихи обыкновенной, тогда как для зерна гречихи татарского — ~ 3.5 мг / кг транс-ресвератрола. Ли и др. [72] сообщили, что ресвератрол в отрубях гречихи Tartary не обнаруживает обнаруживаемого ресвератрола в связанной форме. Chen et al. [73] предложили новую смесь трех растворителей для эффективного и комплексного извлечения фенольных соединений из винной гречихи: ацетон, этилацетат и этанол. Этот метод ценен для оценки функциональных свойств татарской гречки. Экстракция рутина из образцов гречихи более эффективна при использовании 70% этанола вместо более концентрированных экстрактов [74].Поскольку рутин и другие фенольные вещества могут быть связаны с различными соединениями и структурами зерна, эффективная экстракция может занять несколько часов [30,31,59]. Фенольные соединения гречихи могут подавлять развитие грибов из-за фенольных гидрофобных взаимодействий с клеточными мембранами [75]. Этот эффект важен для противогрибковых свойств закваски. Молочнокислые бактерии могут расщеплять флавоноидные гликозиды на флавоноидные агликоны и сахар, а также могут дополнительно метаболизировать агликоны. Образующиеся метаболиты, в состав которых входят молочная кислота и другие органические кислоты, также служат для повышения противогрибковой активности гречневой закваски.Это могло бы объяснить длительный срок хранения хлебобулочных изделий на гречневой закваске по-татарски [75]. Свойства медленного переваривания крахмала были изучены Luo et al. [76], после этанольного экстракта гречихи татарской. Оказалось, что медленная усвояемость этого крахмала связана с воздействием на крахмал фенольных веществ. В своих экспериментах in vivo мыши показали снижение гликемических реакций после приема пищи. Эти данные Luo et al. [76] для зерна гречихи по-татарски и гликемические реакции были аналогичны тем, которые были получены ранее для гречихи обыкновенной [19].Фенольные соединения часто трансформируются в кишечнике до их всасывания. Микробиота кишечника играет важную роль в этом процессе [77]. Диетические фенольные смолы большого размера плохо усваиваются, в то время как продукты микробного преобразования небольшого размера легче всасываются в толстой кишке. Wieslander et al. [78,79] провели комплексное двойное слепое перекрестное исследование с 62 взрослыми участницами женского пола, которые в дополнение к своей обычной диете потребляли либо 359,7 мг рутина в день (диета с высоким содержанием рутина; как гречневое печенье по-татарски), либо 16.5 мг рутина в день (диета с низким содержанием рутина; как обычное гречневое печенье). Через две недели группы изменили свой тип печенья (и, следовательно, уровни потребления рутина) на следующие две недели. Уровни миелопероксидазы (как маркера воспаления) в сыворотке крови значительно снизились у женщин, которые изменили диету с низкого на высокий уровень рутина (неделя 2 по сравнению с неделей 4: снижение на 55,4 мкг / мл; p78). Гречишное печенье также было связано со значительным улучшением симптомов усталости по сравнению с исходным уровнем, в соответствии с уменьшением визуальной аналоговой оценочной шкалы (32 vs.22; p 0,01). Для уровней общего холестерина в сыворотке они были значительно снижены по сравнению с исходным уровнем до четырех недель для комбинации данных для низкого и высокого уровня рутина независимо от порядка приема (5,31 против 4,59 ммоль / л; p80) сообщили, что флавоноиды важны для антигенотоксические эффекты татарской гречихи, хотя другие метаболиты гречихи также имеют важные эффекты. Циклиты (также известные как D-хиро-инозитолы) также были зарегистрированы в зерне татарской гречихи (0,18–0,20%) [81]. Запускается синтез циклитов. параметрами окружающей среды, такими как солевой стресс и засуха, и они могут действовать как криопротекторы.Накопление метаболитов циклита напрямую связано с факторами абиотического стресса. Таким образом, условия окружающей среды растений влияют на регуляцию метаболических путей синтеза и накопления циклитолов [82]. Эти соединения, как известно, обладают антидиабетической, противовоспалительной и другой биоактивностью у людей, как описано Ratiu et al. [82]. Предложены неинвазивные методы различения образцов татарской и гречневой крупы, основанные на накоплении флавонола в зеленых частях растений [83].Здесь пара-анисовая кислота в листьях гречихи указывает на татарскую гречку, а не на обычную гречку. Потребление настоев из татарской гречки — традиция, популярная в Китае, а в последнее время ее потребление распространилось также на Японию и Европу [84]. На модели мышей было сообщено, что экстракты трав в сочетании с настоем из гречневой крупы могут снизить уровень глюкозы в крови и снизить уровень триглицеридов в сыворотке крови, общего холестерина и холестерина липопротеинов высокой плотности. напитков и печенья.Солод богат ориентином, витексином, рутином и кверцетином, хотя уровни флавоноидов в печенье, приготовленном из зерна татарской гречихи, ниже, чем ожидалось, с точки зрения количества сырого зерна татарской гречихи, замоченного или проросшего зерна или солода. Интересно, что уровни этих флавоноидов в татарском гречневом солоде были выше, чем в неповрежденных, вымоченных или проросших продуктах из татарской гречки [37,85]. В сырой татарской гречке оно составляло 2,2 мг / г сухого веса. рутина и цельнотатарского гречишного солода 3.7 мг / г с.в. рутина [37] .Li et al. [72] сообщили об антипролиферативном эффекте фенольных экстрактов татарской гречихи на клетки рака груди человека на клеточной модели in vitro. Эти данные еще предстоит подтвердить экспериментами in vivo. В гречихе, подвергнутой гидротермической обработке, во время замачивания зерна рутин перемещается из фракции отрубей в эндосперм, что приводит к увеличению количества рутина в муке [86]. Подобная миграция более мелких молекул из лузги в крупу при гидротермальной обработке была показана ранее для гречихи обыкновенной [43].Содержание антрахинона в татарской гречке изучалось в зависимости от цвета шелухи зерна [87]. Уровни фагопирина (рис. 3) максимальны во время прорастания семян, а свет важен для превращения протофагопиринов в фагопирины, поскольку было показано, что повышенные уровни фагопирина сопровождают повышенные условия освещения [88,89]. Употребление в пищу зеленых частей гречихи может вызвать фагопиризм, который включает фотосенсибилизацию с серозным экссудатом, раздражение кожи и отек [88,89].У грибов фагопирин участвует в регуляции роста и морфологии мицелия, а также в патогенности [90]. Сообщалось об эмодине (рис. 4) для отрубей и листьев татарской гречихи [91]. По-видимому, он является предшественником в синтезе гиперицина и фагопирина татарской гречихи [92]. Здесь важно то, что эмодин, выделенный из зерна татарской гречихи, стыкуется со всеми тремя активными участками РНК-связывающего домена нуклеокапсидного фосфопротеина SARS-CoV-2 [91,92,93,94].Кверцетин обладает потенциальным терапевтическим действием при остром повреждении почек и для лечения нарушения функции почек. Экстракты зерен гречихи богаты такими флавоноидами, и они могут облегчить вызванное этанолом повреждение печени у крыс [87]. Рутин также защищает мышей с диабетом 2 типа от повреждения печени [95]. Та же функция наблюдается и у экстрактов зерен татарской гречихи за счет ингибирования гибели митохондриальных клеток [96,97]. Фармакокинетика in vivo также подтвердила назначение экстрактов татарской гречихи для профилактики алкогольной болезни печени у людей [98].Сообщается, что рутин также эффективен при заживлении ран и у крыс с гипергликемией, поскольку он может снижать окислительный стресс и воспалительные реакции, тем самым снижая риск образования язвы [99]. Рутин индуцирует активность глутатионпероксидазы, которая защищает семенники взрослых крыс от воздействия этанола [100]. Флавоноиды татарской гречихи улучшают чувствительность сосудов и проявляют антигипертензивный эффект у крыс со спонтанной гипертонией [101]. Салицилальдегид является наиболее характерным соединением для гречихи обыкновенной, и в татарской гречке он не обнаружен.Аромат татарской гречки существенно отличается от аромата гречки обыкновенной; действительно, поскольку салицилальдегид является летучим соединением, его можно использовать в качестве маркера для обнаружения загрязнения татарской гречки гречихой обыкновенной [43,44]. Гречка по-винному также содержит нафталин [44].6. Выводы
Продукты, приготовленные из зерна татарской гречихи, показали профилактический эффект против ряда хронических заболеваний, включая ожирение, сердечно-сосудистые заболевания, образование камней в желчном пузыре и гипертонию.Эффект в основном объясняется устойчивым крахмалом, белком и фенольными веществами в зерне, а также взаимодействиями между этими составляющими. Полифенолы влияют на усвояемость белков после гидротермальной обработки. Их взаимодействие снижает переваривание белка через тонкий и толстый кишечник. Микробные процессы в толстой кишке повышают усвояемость зернового протеина и крахмала, которые в противном случае блокируются полифенолами в гречихе, подвергнутой гидротермической обработке.Среди полифенолов фагопирин, по-видимому, представляет угрозу для здоровья при употреблении зеленых частей растений, особенно летом, людьми со светлой кожей. Доказано, что потребление гречневого зерна и продуктов из него безопасно.
Гречневая крупа может снижать уровень холестерина в сыворотке крови за счет увеличения экскреции стероидов с калом, которая вызвана связыванием стероидов с непереваренным белком. Устойчивые к пищеварению пептиды в значительной степени ответственны за выведение желчных кислот.Поскольку гречка не содержит белков глютена, ее используют в пищу людям с глютеновой болезнью. Сбалансированный аминокислотный состав белков гречки представляет собой важный источник диетического белка для людей, придерживающихся вегетарианской или веганской диеты. Повышение содержания стойкого к перевариванию крахмала достигается при варке гречневой крупы, а также обработкой холодной плазмой и комплексообразованием кверцетина, что приводит к модификации крахмала из татарского зерна гречихи. Резистентный крахмал действует как пищевое волокно и выполняет функцию пребиотика.Гречиха по-татарски — это культура, которая традиционно используется в пищу в горных районах Азии и Европы. По своему происхождению гречиха татарская — растение с низким потреблением энергии. Благодаря содержанию флавоноидов и других фенольных веществ гречиха татарская устойчива к болезням растений, вредителям и повреждениям УФ-В излучением. Это позволяет выращивать татарскую гречку как органическую и экологическую культуру без необходимости добавления искусственных удобрений или химической обработки.
И последнее, но не менее важное: татарская гречка напоминает многим потребителям о «старых добрых временах», когда популярность татарских гречневых блюд растет, особенно среди людей, заботящихся о качестве продуктов питания, в Азии и Европе.Это дает возможность разрабатывать новые, более «современные» продукты питания, основанные на старых кулинарных традициях, с переоценкой на основе современных научных знаний качества и потенциала татарской гречки.
Разработка системы штрих-кодов на основе 50 InDel для генетической идентификации ресурсов татарской гречихи
Abstract
Гречиха татарская ( Fagopyrum tataricum Gartn.) — высокофункциональная культура, которая может стать целью многих будущих селекционных работ.Надежное сохранение различных генетических ресурсов ex situ имеет важное значение для современного селекции винных сортов гречихи. Мы разработали кодоминантные инсерции / делеции (InDel) маркеры на основе ПЦР для различения генетических ресурсов винной гречихи. Во-первых, мы получили полный геном из 26 образцов в эталонном геноме суперкаффолда размером 569,37 МБ для винной гречихи cv. «Тэгван 3–7». Затем 171 926 гомогенных и 53 755 гетерогенных InDel были обнаружены путем сравнения 26 образцов с эталонной последовательностью «Daegwan 3–7».Из них 100 кандидатов InDel длиной от 5 до 20 п.н. были выбраны для валидации, и 50 из них выявили полиморфизм между 26 образцами и «Daegwan 3-7». Утвержденные InDels были дополнительно протестированы путем оценки их вероятности дать начало одному или нескольким ПЦР-продуктам в 50 других образцах, охватывающих большинство типов генома винной гречихи. Частоты основных аллелей варьировались от 0,5616 в локусе TB42 до 0,9863 в локусе TB48 со средним значением PIC 0.1532 в диапазоне 0,0267–0,3712. Чтобы создать удобную для пользователя систему, гомология генотипов между образцами и между образцами была визуализирована как в одном (1D), так и в двухмерном (2D) типах штрих-кода путем сравнения полиморфизмов ампликонов с эталонным сортом, «Дэгван 3–7». . » Филогенетическое дерево и популяционная структура 76 образцов в соответствии с полиморфизмом ампликона для 50 маркеров InDel соответствовали образцам, использующим несинонимичные варианты однонуклеотидного полиморфизма, что указывает на то, что система штрих-кодов, основанная на 50 InDel, была полезным инструментом для повышения надежности выявление образцов винной гречихи в запасах зародышевой плазмы.
Образец цитирования: Sohn H-B, Kim S-J, Hong S-Y, Park S-G, Oh D-H, Lee S и др. (2021) Разработка системы штрих-кодов на основе 50 InDel для генетической идентификации ресурсов татарской гречихи. PLoS ONE 16 (6): e0250786. https://doi.org/10.1371/journal.pone.0250786
Редактор: Химаншу Шарма, Национальный институт агропродовольственной биотехнологии (НАБИ) Мохали, ИНДИЯ
Поступила: 28 сентября 2020 г .; Принята к печати: 14 апреля 2021 г .; Опубликовано: 3 июня 2021 г.
Авторские права: © 2021 Sohn et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией.
Финансирование: Это исследование финансировалось грантами Программы совместных исследований для развития сельскохозяйственных наук и технологий, финансируемой Управлением развития сельских районов (RDA) Республики Корея для YHK (Проект №PJ01189401) и TheragenEtex Inc. предоставили поддержку этому исследованию в виде заработной платы SGK. Конкретная роль этих авторов сформулирована в разделе «Авторский вклад». RDA оценивало проект каждый год в течение 5 лет (2016-2020 гг.) И принимало участие в принятии решения об отправке для публикации, но спонсоры не играли дополнительной роли в дизайне исследования, сборе и анализе данных или подготовке рукописи.
Конкурирующие интересы: Авторы ознакомились с политикой журнала, и у авторов этой рукописи есть следующие конкурирующие интересы: SGK является оплачиваемым сотрудником TheragenEtex Inc.и SL входит в совет директоров EONE DIAGNOMICS Genome Center Co. Ltd. Это не влияет на нашу приверженность политике PLOS ONE в отношении обмена данными и материалами. Нет никаких патентов, продуктов в разработке или продаваемых продуктов, которые можно было бы декларировать.
Введение
Семейство эвдикотовых Polygonaceae состоит из 27 зарегистрированных видов, включая 2 культивируемых вида, гречиху обыкновенную ( Fagopyrum esculentum Moench) и татарскую гречку ( F . tataricum Gaert.) [1,2]. Гречиха ( F . esculentum и F . tataricum ) культивируется как короткосезонный псевдозерновой во многих странах и регионах, включая Китай, Непал, Россию, Европу, Корею и Японию [3 , 4]. После одомашнивания на возвышенностях юго-западного Китая древние фермеры создали множество местных сортов татарской гречихи путем искусственного и естественного отбора [5]. После выращивания в Китае эти староместные сорта сначала распространились в Непал, Россию, Европу, Корею и Японию [5–9].Некоторые характеристики винной гречихи, такие как ее самоопыляемая природа и способность адаптироваться к различным регионам (от высокогорных зон до низин), привели как к морфологии, так и к генетически дивергентным очагам староместных сортов [10–13]. Эти староместные сорта были в значительной степени заменены меньшим количеством староместных сортов с особыми желательными характеристиками, такими как высокая урожайность, засухоустойчивость и аромат [5,14].
Гречиха татарская является основной пищевой культурой в высокогорных зонах (например, в высокогорных районах южного Китая и в Гималайских холмах) из-за ее морозостойкости в плохих почвенных условиях.В последнее время гречиха также используется для различных целей, от различных продуктов переходного периода до медицинских [15,16]. Зерно винной гречки считается идеальным источником функциональной пищи для человека, поскольку оно более богато белками, жирами, витаминами, рутином, кверцетином и другими флавоноидами, чем обычная гречка [17–23]. Популярность татарской гречки неуклонно растет, и ее выращивание распространилось на районы, находящиеся далеко за пределами своего первоначального региона [10,11,24,25].Это свидетельство адаптируемости татарской гречки, которая будет иметь решающее значение в условиях меняющегося климата Земли [14]. Селекция товарных сортов татарской гречихи улучшает сельскохозяйственные характеристики, такие как полегание, поздняя спелость и низкая урожайность [26,27]; однако это культивирование явилось результатом узкой генетической базы только нескольких сортов, из которых были получены современные сорта, которые плохо приспособлены к различным условиям выращивания. Узкое генетическое разнообразие коммерческого сорта обусловлено жестким качеством, требуемым фермерами и переработчиками, ограниченным использованием экзотической зародышевой плазмы, ограниченными стратегиями селекции и индивидуальным отбором растений [28–30].Таким образом, большее генетическое разнообразие генотипов винной гречки может быть использовано для улучшения конкретного представляющего интерес признака из-за узкого генетического разнообразия современных коммерческих сортов [14,31,32].
Идентификация зародышевой плазмы винной гречихи имеет жизненно важное значение для увеличения генетического разнообразия. Исследователи были сосредоточены на разработке молекулярных маркеров с использованием зародышевой плазмы и морфологических дескрипторов, чтобы обеспечить высокую способность распознавания. Существует множество методов различения разновидностей на основе полиморфизма ДНК, таких как случайная амплификация полиморфной ДНК (RAPD), полиморфизм длины амплифицированного фрагмента (AFLP), межпростой повтор последовательности (ISSR), простые повторы последовательности (SSR) и одиночный нуклеотид. полиморфизм (SNP).Полиморфизмы вставки-делеции (InDels) привлекают все больше внимания ученых, занимающихся молекулярной селекцией, поскольку они просты в использовании, совмещены (полностью информативны) и относительно многочисленны [33-40]. В гречневой каше SSR [9], AFLP [41], RAPD [42,43], ISSR [44] и SNP [45,46] использовались для оценки генетической изменчивости, но ни одно исследование не использовало различные методы дискриминации. с маркерами InDel из-за сложности обнаружения полиморфизмов ДНК в образцах винной гречихи.
До недавнего времени отсутствие крупномасштабной информации о секвенировании генома привело к внедрению InDels в качестве генетических маркеров у немодельных видов (например,г., гречка обыкновенная и винная) сложно. В последние годы опубликованные исследования предоставили проекты геномов гречихи обыкновенной [47] и сорта гречихи винного сорта Pinku1 [48] в семействе Polygonaceae. Что еще более важно, огромное количество данных о полиморфизме теперь может быть получено из огромного количества данных секвенирования с использованием высокопроизводительных технологий секвенирования [49]. Кроме того, Sohn et al. [38] сообщили, что система штрих-кодов на основе InDel ориентирована на удобство использования и обеспечивает эффективную систему управления ресурсами.В дополнение к системе штрих-кодов на основе InDel в настоящее время появляются морфологические дескрипторы, предназначенные для идентификации образцов татарской гречихи, которые могут использоваться для проверки ключевых геномных факторов, которые объясняют или предсказывают основные агрономические признаки.
Мы представляем высококачественный черновой вариант сборки генома и аннотацию винной гречихи. Ф . tataricum cv. «Тэгван 3–7». Используя подходы сравнительной геномики, мы выявили 171 926 гомогенных InDel и 53 755 гетерогенных InDels, обнаруженных в 26 образцах винной гречихи, которые мы сравнили с черновыми геномами сорта винной гречихи «Daegwan 3–7.Среди них 50 полиморфных InDel из 26 образцов были отобраны с помощью гель-электрофореза, которые были преобразованы в штрих-коды путем сравнения полиморфизмов ампликонов с эталонной последовательностью. Мы также выявили географическое распределение 73 образцов путем анализа структуры населения с использованием штрих-кодов. Для генетической идентификации ресурсов винной гречки мы создали удобную систему штрих-кодов на основе 50 генотипов 73 образцов на основе InDel. В качестве удобной для пользователя системы гомологию между образцами можно прогнозировать как в одном (1D), так и в двухмерном (2D) в виде блоков.Наша платформа может быть использована в программах генетических исследований и селекции, а также для эффективных систем управления ресурсами гречихи винного сорта.
Материалы и методы
Растительные материалы
Образцы 73 растений татарской гречихи (таблица S1) были предоставлены Национальным центром агробиоразнообразия (Чонджу, Республика Корея) и Научно-исследовательским институтом сельского хозяйства Хайленд (Пхенчхан, Республика Корея). Образцы были посажены в теплице, расположенной в Научно-исследовательском институте сельского хозяйства Хайленд (37 ° 68 ′ с.ш. и 128 ° 73 ′ в.д., высота 779 м), провинция Канвон, Республика Корея.Эти растения самоопылялись 3 раза методом опускания одного семени (SSD) путем выращивания отдельных растений в цветочных горшках (диаметром 10 см) [50]. Мы вырастили еще четыре поколения HLB1001 с использованием метода SSD, что привело к отбору потомства «Daegwan 3–7» для чернового генома татарской гречихи. Всего было собрано 26 образцов (Китай, Индия, Непал, Бутан, Пакистан и США).
Для изучения биоразнообразия и сравнительных взаимосвязей между образцами собранные образцы были посеяны в 2017 году и выращены в теплице в Научно-исследовательском институте сельского хозяйства Хайленд.Мы разделили растения по стадиям развития: стадия вегетативного роста (1–40 дней после посадки, DAP), стадия цветения (40–80 DAP) и стадия урожая (80–90 DAP). На стадии урожая цвет кожуры семян стал черным, что указывало на полное созревание семян [51].
Измерение содержания рутина и кверцетина
Семена стадии урожая использовали для измерения содержания в них рутина и кверцетина. Сначала зрелые семена 76 образцов измельчали до мелкого порошка, затем образцы немедленно замораживали в жидком азоте и хранили при -80 ° C до дальнейшего использования.Всего 0,10 г сухого порошка для каждого образца смешивали с 1 мл метанола, затем экстрагировали при 80 ° C в течение 1 часа в обычном аппарате Сокслета. Экстракт фильтровали через шприцевой фильтр 0,20 мкм (PTFE 13 мм, PALL Life Sciences, Анн-Арбор, Мичиган) на содержание рутина [23]. Ультраэффективную жидкостную хроматографию (Waters Corporation, Милфорд, Массачусетс, США) проводили на Acquity 1-класса с использованием колонки C18 (2,1 мм × 100 мм, 1,7 мкм) (Waters Corporation, США) при 30 ° C. Подвижная фаза (1% муравьиная кислота в воде / 0.1% муравьиной кислоты в ацетонитриле) пропускали методом градиентного элюирования (таблица S2) со скоростью 0,25 мл / мин с общим объемом впрыска 10 л. Количество рутина оценивали на основании линейной калибровочной кривой для стандартного рутина и кверцетина (Extrasynthese, Франция) при длине волны детектирования 259 нм. Для каждого образца было выполнено три независимых анализа образцов. Все статистические анализы были выполнены с использованием вер. 3.6.1 программного обеспечения R.
секвенирование ДНК и
de novo сборкаПосле 21 DAP листья в третьем узле потомства «Daegwan 3–7» использовали для создания чернового генома.Сначала были собраны трехнедельные листья основной массы 10 растений для экстракции геномной ДНК с использованием стандартного протокола CTAB (цетилтриметиламмонийбромид) [52,53]. Библиотеки как короткого чтения (Illumina), так и библиотеки длительного чтения (PacBio) были подготовлены в соответствии с инструкциями производителя для всего генома F . tataricum . Библиотеку с короткими вставками с парными концами (PE) получали с использованием набора для подготовки образцов ДНК Illumina TruSeq Nano (Illumina Inc., Сан-Диего, Калифорния, США) и 1 ~ 3 мкл ДНК.Анализ профиля библиотеки выполняли с помощью биоанализатора Agilent 2100 (Agilent Technologies, США) и количественного анализа qPCR, затем библиотеки секвенировали до 2 × 100 п.н. на платформе Illumina HiSeq 2500 (Illumina, США) от TheragenEtex Bio Institute (TheragenEtex Inc. ., Сувон, Республика Корея). Библиотеки PacBio SMRTbell (вставки 2 кб, 5 кб, 10 кб и 15 кб) были приготовлены с использованием стандартных протоколов подготовки библиотеки PacBio, которые доступны на http://pacificbiosciences.com/. Секвенирование проводили на системе PacBio RS II (Pacific Biosciences, США) с использованием химии C4 и 240-минутных фильмов TheragenEtex Bio Institute.
Короткие и длинные чтения были собраны отдельно. Сборка de novo длинных чтений PacBio была выполнена с использованием Fast Alignment и CONsensus [54] с параметрами по умолчанию. Короткие чтения были собраны с помощью SOAPdenovo2 [55] с параметрами по умолчанию. Исходные контиги были объединены с помощью HaploMerger2 [56]. Затем были использованы как короткие, так и длинные чтения для конструирования каркасов с SSPACE [57], что привело к геному чернового варианта суперкафолдной крупы винной гречихи cv. «Тэгван 3–7». Оценивали длину N50 конечных сборок генома.
Идентификация и проверка полиморфных InDels
Геномная ДНК была извлечена для повторного секвенирования всего генома из 26 образцов, которые 3 раза самоопылялись методом SSD. Трехнедельные листья из основной массы 10 растений были собраны для экстракции геномной ДНК с использованием стандартного протокола CTAB [52,53]. Данные состояли из считываний размером 101 п.н., которые были получены с использованием секвенатора HiSeq 2500. Программа 2 короткого олигонуклеотидного выравнивания (SOAP2) была использована для картирования необработанных считываний парных концов на эталонный геном (гречиха винная сорт cv.«Тэгван 3–7»). Для каждого образца более 93% считываний были правильно сопоставлены с эталонным геномом. Размер вставки оценивался путем сопоставления считываний с эталонным геномом с использованием алгоритма выравнивания Берроуза-Уиллера (bwa) [58] версии 0.5.9. Выровненные чтения были повторно выровнены в положениях InDel с использованием алгоритма GATK InDelRealinger [59] для повышения качества отображения. Базовые показатели качества были перекалиброваны с использованием алгоритма GATK TableRecalibration. Пара праймеров, использованная для амплификации каждого из InDel, была выбрана с помощью программного обеспечения Primer3 (http: // primer3.sourceforge.net). Мы выбрали InDel с более чем 10 п.н. и разработали праймеры в соответствии с характеристиками каждого InDel с использованием программного обеспечения Primer 3.
Анализ полиморфизма ампликона и процесс штрих-кодирования
Анализ ПЦРвыполняли с использованием 10 мкл реакционных смесей, содержащих 20 нг общей геномной ДНК, 2 мкМ праймера и 5 мкл смеси GoTaq Green Master Mix (Promega, Мэдисон, Висконсин, США). ПЦР выполняли при 95 ° C в течение 5 минут с последующими 35 последовательными циклами при 94 ° C в течение 30 секунд, 45 ° C в течение 30 секунд и 72 ° C в течение 30 секунд с использованием Mastercycler pro 384 (Eppendorf, Германия).Продукты ПЦР разделяли электрофорезом в 3% гелях сертифицированной ультраагарозы низкого диапазона (Bio-rad) с последующим окрашиванием LoadingSTAR (Dynebio, Республика Корея). Для ограничения количества кандидатов мы выбрали пары праймеров, которые амплифицировали продукты ПЦР длиной 150–231 п.н. Аллельное разнообразие InDel с продуктами е-ПЦР в 26 геномах винной гречки оценивали с помощью PIC (Polymorphism Information Content), который был определен как PIC, где p ij — частота j -го паттерна. для маркера и [60,61].
50 маркеров InDel были отобраны в соответствии с успешностью их генотипирования и размером полосы ПЦР. Различительная способность отобранного набора 50 InDel для идентификации винной гречки оценивалась с использованием 76 образцов винной гречки. Праймеры были сконструированы путем нацеливания на область InDel таким образом, чтобы генотипы образцов винной гречихи давали одинаковые или разные (вставки или делеции) ампликоны относительно эталонного генома, «Дэгван 3–7». На основании результатов амплификации маркера InDel тот же результат с эталонным геномом был представлен буквой «а», а другие результаты были представлены буквой «b».Для представления результатов с помощью штрихового кодирования «a» и «b» были преобразованы в белый и черный цвета соответственно. Гомологию образцов рассчитывали после ПЦР-амплификации всех 50 InDel.
Филогенетический анализ и популяционная структура 73 образцов татарской гречихи
Различительная способность отобранного набора 50 InDel для идентификации винной гречихи была оценена с использованием 76 образцов винной гречки. Анализ проводился с помощью программы POWERMARKER Ver.3.23 (Лю и Муза 2005; http://www.poxermarker.net). Филогенетическое дерево для 73 образцов было составлено на основе генотипов, определенных с помощью 50 InDel, с использованием метода взвешенного соединения соседей с простыми коэффициентами соответствия в программе DARwin [55]. Для определения кластеров индивидуумов на основе их генотипов использовалась основанная на модели СТРУКТУРА 2.3.4 [63] с моделью примеси, периодом выгорания в 100 000 итераций и данными из 50 локусов InDel. Количество кластеров (k) было установлено равным пяти, поскольку это количество максимизировало параметр Δk (специальный критерий).Мы использовали моделирование со значениями k в диапазоне от 2 до 10 с 5 повторениями, чтобы вычислить значение LnP (D). Оптимальное значение k можно выбрать на основе максимального LnP (D), который обеспечивает четкую структуру популяции. Исходя из этого, мы выбрали K = 3 (три группы предков) для популяций винной гречихи. Чтобы проверить качество этих анализов, были составлены филогенетическое дерево и популяционная структура для 26 повторно секвенированных образцов на основе генотипов, определенных с использованием 6622 несинонимичных SNP.
Результаты
Сборка генома и полногеномное секвенирование образцов татарской гречихи
Мы вырастили семь поколений сорта гречиха татарская. «Дэгван» с использованием метода SSD и изолированного человека (обозначенного как «Дэгван 3–7»), чтобы исключить любую возможную неоднородность в исходной партии семян. Мы произвели в общей сложности 43,83 и 32,17 ГБ последовательностей с платформ секвенирования Illumina PE и Single-Molecule Real-Time (SMRT) соответственно, что соответствовало 78x (Illumina PE, таблица S2) и 58x (SMRT, таблица S3) (S1 Рис) для эталонного генома.Комбинируя стратегии секвенирования PacBio и Illumina Hiseq2500, мы получили окончательный проект сборки 569,37 МБ в 2886 каркасов с 50% общей последовательности, захваченной в 156 каркасов больше 886968 bps (Таблица 1, N50).
Нами было отобрано 26 образцов татарской гречихи для полногеномного ресеквенирования. Эти образцы возникли или были популяризированы в различных азиатских и международных регионах, включая 15 местных сортов из Китая, 4 из Непала, 4 из Индии, 1 из США, 1 из Бутана и 1 из Пакистана (Таблица 2).После того, как 3 дополнительных поколения были выращены с использованием метода SSD, панель из 26 генотипов показала широкую степень вариации содержания рутина и фенотипа (рис. 1 и 2). 26 образцов показали различное распределение содержания рутина по сравнению с 73 образцами (рис. 2). В частности, у HLB1013 было самое высокое содержание рутина (1 921 мг / 100 г · DW), а у HLB1006 было самое низкое (1250 мг / 100 г · DW) среди 26 образцов. Были обнаружены большие различия в форме семян и цвете кожуры между 26 образцами (рис. 1).Информация о семенах соответствовала известной генетической изменчивости 26 образцов.
Рис. 2. Содержание рутина в 73 образцах татарской гречихи в зависимости от страны происхождения.
Фенотипическая сегрегация показана в формате прямоугольной диаграммы. Межквартильный регион, медиана и диапазон обозначены прямоугольником, жирной горизонтальной линией и вертикальной линией соответственно. США / Япония / Россия включает 4 присоединения; 1 из США, 1 из Японии и 2 из России. Мы отобрали 26 образцов для повторного секвенирования всего генома, которые возникли или были популяризированы в различных азиатских и международных регионах.Более подробную информацию о 73 образцах татарской гречки см. В таблице S1.
https://doi.org/10.1371/journal.pone.0250786.g002
Мы произвели считывания парных концов ДНК на глубине 23–27 раз (таблица 2) для 26 геномов образцов и сопоставили их с эталонным геномом ( «Тэгван 3–7»). Качество секвенирования всех образцов было высоким: 93–95% считываний образцов доступа были сопоставлены с эталонным геномом, а 96–97% эталонного генома были покрыты (таблица S3).Генетические вариации (SNP и небольшие индексы) среди 26 образцов, собранных из шести стран, были индивидуально определены Samtools и Genome Analysis Tollkit GATK (см. Методы). Для повышения точности прогноза для последующего анализа использовались только вариации, предсказанные обоими методами. Биоинформатический анализ выявил 171 926 и 53 755 гомогенных и гетерогенных InDel соответственно (рис. 3). Гетерозиготность всех образцов составляла около 10%, что отражает возможность перекрестного опыления, хотя использовалось самоопыление (таблица S4).
Рис. 3. Количество InDels в 26 образцах татарской гречихи, выровненных с эталонным геномом «Дэгван 3–7».
Количество вариантов, однородных альтернативному генотипу; оба аллеля совпадают с альтернативным генотипом. Количество вариантов гетерогенного генотипа; один аллель соответствует эталонному, а другой — альтернативному генотипу.
https://doi.org/10.1371/journal.pone.0250786.g003
Выбор 50 инделов и построение базы данных по 73 образцам штрих-кодов
Сравнение последовательностей на нуклеотидном уровне тестируемых геномов с эталонным геномом выявило InDels, которые можно легко использовать для идентификации ресурсов винной гречки в обычных лабораториях.В соответствии с тем, что было обнаружено у других организмов [38,64], подавляющее большинство гомогенных InDel из всех образцов были короткими с преобладанием 4-bp или меньше событий (Fig 3). Из этих данных полиморфизма мы выбрали InDels с более чем 10 пар оснований для дальнейшего анализа, чтобы помочь нам идентифицировать их с помощью ПЦР и гель-электрофореза. Мы протестировали 100 наборов праймеров, сосредоточив внимание на их способности амплифицировать продукты ПЦР в 26 типах генома, поскольку это исследование было направлено на установление основанных на ПЦР маркеров, применимых для всех образцов татарской гречихи.Праймеры для ПЦР были разработаны для генерации ампликонов различного размера в интервале 150–231 п.н. (таблица S5). Среди них 50 локусов InDel были широко распределены на целых хромосомах в эталонном геноме («Pinku 1»), о котором сообщалось ранее (рис. 4). Мы выбрали 50 InDels для проверки точности прогноза и обнаружили, что 98,2% результатов ПЦР в 26 образцах соответствовали прогнозируемым результатам. Результаты анализа InDel были очень точными (таблицы S5 и S6). Эта высокая точность обеспечила прочную основу для нашего анализа данных и предоставляет данные высокого качества для будущего анализа данных.
Праймерыбыли сконструированы путем нацеливания на область InDel таким образом, чтобы генотипы 26 образцов производили одинаковые или разные (вставки или делеции) ампликоны относительно эталонного генома (рис. 5). При использовании ранее описанного метода штрих-кода [38,39], результаты, идентичные референсному геному «Дэгван 3–7», были представлены буквой «а», тогда как другие результаты были представлены буквой «б», которые были представлены как «белый» »И« черный »штрих-коды соответственно. На рис. 5 показана система штрих-кода, примененная к образцам татарской гречихи HLB1007 и HLB1008, которые были преобразованы в стандартные одномерные и широко используемые двухмерные штрих-коды путем сравнения их полиморфизмов ампликонов с эталонным геномом «Дэгван 3–7.Этот результат подчеркивает, что типы штрих-кодов могут быть эффективно использованы для исследования степени маркерных различий в образцах винной гречихи (рис. 6). Создание, стабильность и качество системы штрих-кодов для генетической идентификации с использованием отобранных 50 InDel тщательно оцениваются ниже.
Рис. 5. Применение системы штрих-кодов винного камня для присоединения винного камня, HLB1007 и HLB1008.
(A) Продукты амплификации и штрих-кодовое представление полиморфизмов с использованием маркера InDel TB11 на образцах винной гречихи.Результаты ПЦР с использованием 50 InDel в (B) HLB1007 и (C) HLB1008 были преобразованы в стандартные 1D и широко используемые типы 2D штрих-кодов по сравнению с таковыми в «Daegwan 3–7». Результаты, идентичные эталонному геному, были представлены белым цветом, а другие результаты — черным.
https://doi.org/10.1371/journal.pone.0250786.g005
Рис. 6. Сравнение двухмерных штрих-кодов между HLB1007 и HLB1008.
Идентичные результаты отображаются белым цветом, а другие результаты — черным.Морфология семян HLB1007 и HLB1008.
https://doi.org/10.1371/journal.pone.0250786.g006
Создав базу данных 50 полиморфизмов InDel для 73 образцов татарской гречки (рис. S2), мы создали более стабильную основу для использования штрих-кода. Система представляет собой многообещающий инструмент для определения генетических ресурсов винной гречихи. Частоты аллелей и среднее значение PIC для 50 локусов InDel показаны в таблице 1. Частоты основных аллелей варьировались от 0.5616 в локусе TB42 до 0,9863 в локусе TB48. Среднее значение PIC составило 0,1532 с диапазоном 0,0267–0,3712, что означает, что выбранные InDels могут быть применены для исследования полиморфизма образцов винной гречихи (таблица S7). Генотипы 73 образцов татарской гречихи были сконструированы с помощью двумерного штрих-кода, и генотипическая дискриминация была проанализирована путем сравнения его с образцами системы штрих-кода. Средняя разница между проанализированными образцами составила 7 InDels из 50 InDels, в то время как 6 образцов имели различия в 12 InDels по сравнению с эталонным геномом «Daegwan 3–7.Сравнивая двумерные штрих-коды образцов, можно было легко визуально различить различные блоки, чтобы можно было определить различия генотипов образцов (рис. 6). Различительная способность генетических ресурсов была достаточно высока, чтобы расширить наше понимание как филогенетических сигналов, так и изменчивости на уровне популяции.
Филогенетический анализ и популяционная структура образцов татарской гречихи
Филогенетический анализ 6622 несинонимичных вариантов SNP и популяционная структура разделили панель на 2 первичные популяции (рис. 7): контрольную группу и не контрольную группу.Аналогичный результат был достигнут с данными SSR, которые указали на два отдельных групповых кластера винной гречки [9]. 23 образца имели 4 подгруппы в нереферентной группе и были классифицированы в соответствии с областью сбора. Филогенез решил, что подгруппа 2 (3 Непал, 1 Бутан и 2 Китай) ближе к подгруппе 1 (8 Китая и 1 Индия), чем подгруппа 3 (3 Индия, 1 Пакистан и 1 Непал).
Рис. 7. Анализ разнообразия 26 повторно секвенированных образцов винной гречихи.
(A) Филогенетический анализ на основе несинонимичных SNP.(B) СТРУКТУРА [63] классификация и (C) анализ главных компонентов были аналогичны. Каждый цвет обозначает страну происхождения.
https://doi.org/10.1371/journal.pone.0250786.g007
Для проверки надежности 50 маркеров InDel для различения образцов татарской гречихи были построены бин-карты для 73 образцов татарской гречихи с использованием InDels (таблица S6 ). Мы проанализировали популяционные структуры исследуемых образцов и референтных групп.Результат показан на рис. 8. При K = 3 кластеры были привязаны к Юго-Восточной Азии и Китаю. Среди подгруппы 1 образцы в Юго-Восточной Азии почти полностью состояли из красного компонента, в то время как образцы из Китая состояли из красного, зеленого и синего компонентов. В подгруппу 1 в основном входили образцы из Юго-Восточной Азии (65,4%), в подгруппу 2 — образцы из Китая (71,4%), а в подгруппу 3 — образцы из Китая (97,0%). В то же время китайские образцы подгруппы 1 и 3 показали смешанный состав как красного, зеленого, так и синего компонентов.PCA также дал аналогичные результаты, когда образцы образуют плотный кластер, разделенный областью сбора.
Рис. 8. Анализ разнообразия образцов татарской гречихи.
(A) СТРУКТУРА [63] результаты основаны на 50 InDels: 73 образца были разделены на 3 субпопуляции. (B) Анализ главных компонентов и классификация СТРУКТУРЫ были аналогичны. Каждый цвет обозначает страну происхождения. (C) Филогенетический анализ 73 образцов татарской гречихи с использованием взвешенного анализа слиянием соседей в программе DARwin [62].Горизонтальная полоса указывает расстояние на основе простого коэффициента соответствия. Каждая подгруппа представляет собой субпопуляцию на основе результатов СТРУКТУРА.
https://doi.org/10.1371/journal.pone.0250786.g008
Взвешенные отношения объединения соседей (дерево NJ) с картами ячеек выявили 3 группы (G1, G2 и G3), содержащие 40, 16 и 17 присоединения соответственно (рис. 8). Образцы, собранные в Непале и Бутане, продемонстрировали субструктуру внутри группы 1, являясь генотипами, наиболее близкими к контрольной группе.Другой результат показал, что две отдельные группы были сгруппированы с использованием несинонимичных вариантов SNP (рис. 7). Цветовые обозначения ветвей подтверждают классификацию 73 образцов по регионам сбора (рис. 8). В филогенетическом анализе PCA и популяционная структура показали, что образцы винной гречихи разделились на разные группы, что отражало их географическое распространение. С доказательствами, связывающими 50 InDels с экологической адаптивностью, эти результаты согласуются с разделением татарской гречихи на различные группы в соответствии с их географическим распределением.Эти результаты показывают практическую применимость системы штрих-кодов, которую можно использовать для идентификации генетической изменчивости, связанной с фенотипической изменчивостью, имеющей функциональное значение, а также маркеры, которые можно использовать в селекции сельскохозяйственных культур.
Обсуждение
Татарская гречиха — важная культура, которая может стать целью многих будущих селекционных усилий. Это вид сельскохозяйственных культур, представляющих интерес по разным причинам, включая более высокий уровень рутина по сравнению с гречкой [10,23,65].Для выведения товарных сортов татарской гречки необходимо расширить элитную зародышевую плазму за счет добавления различных генетических ресурсов. Эта область все еще недостаточно представлена на генетическом уровне, несмотря на огромное разнообразие зародышевой плазмы и ее важность в ресурсах, адаптированных к стрессу [66–70]. В частности, поскольку эти ресурсы обладают многими отличительными чертами, мы использовали 73 генетических ресурса винной гречихи для создания основы для сравнительного анализа. Эти ресурсы различаются по морфологии семян (рис. 1) и содержанию рутина (рис. 2).Эти различия могут привести к разным ответам на УФ-В, засуху [69] и солевой стресс [67]. Недавняя публикация геномов винной гречихи предоставила эталонный геном, необходимый для изучения молекулярных маркеров, позволяющих различать эти генетические ресурсы [48]. Узкое генетическое разнообразие коммерческого сорта имеет различные причины, в том числе жесткое качество, требуемое фермерами и переработчиками [28,30]. Таким образом, молекулярные маркеры необходимы для понимания генетических различий образцов винной гречихи, которые могут быть использованы для расширения генетических ресурсов программ коммерческой селекции.Чтобы решить эту проблему, мы создали удобную для пользователя систему штрих-кодов, используя 50 генотипов на основе InDel 73 образцов. Наша система штрих-кодов может использоваться в генетических исследованиях, селекционных программах и эффективных системах управления ресурсами гречихи.
Технологии высокопроизводительного секвенирования [49] предлагают огромные возможности для создания огромного количества данных секвенирования из до сих пор не охарактеризованных геномов, что облегчает внедрение InDels в качестве генетических маркеров в немодельных культурах, таких как татарская гречиха.Учитывая, что существующий эталонный геном для «Pinku 1» был из Китая [48], мы сконструировали новый эталонный геном в масштабе суперкаффолда для различных «Дэгван 3–7» из Республики Корея, объединив стратегии секвенирования PacBio и Illumina Hiseq2500. Используя этот комплексный подход, мы получили размер каркаса N50 463 432 п.н. для чернового генома «Дэгвана 3–7» (Таблица 1). Чтобы изучить геномные вариации образцов татарской гречихи, мы выполнили полногеномное секвенирование 26 образцов в эталонном геноме («Дэгван 3–7») (Таблица 2).Примечательно, что глубина (23 × — 27 ×) охвата последовательностей указывает на то, что она хорошо подходит для анализа последовательностей 26 генетических ресурсов. Из этих сравнений геномов было получено 171926 гомогенных InDel и 53 755 гетерогенных InDel с плотностью InDel 2 на каждый 1 kb посредством попарного сравнения с эталонным геномом (рис. 3 и таблица S5). Среди них 50 полиморфных InDel были отобраны с помощью гель-электрофореза, результаты которого на 98,2% совпали с предсказанными результатами (таблицы S6 и S7).Более того, 49 InDel были широко распространены во всем геноме «Pinku 1». Учитывая, что локус TB8 не был обнаружен в «Pinku 1», два эталонных генома для «Pinku 1» и «Daegwan 3–7» могут служить основой и ссылками для разработки пангенома для уменьшения систематической ошибки ссылки. Планируем и дальше улучшать проект генома гречихи винной сорт. «Дэгван 3–7» для использования в пангеноме. Наши результаты показывают, что черновые варианты геномов de novo суперкафолда обычно облегчают внедрение InDels в качестве генетических маркеров в немодельные культуры, такие как татарская гречиха.
Чтобы эффективно идентифицировать образцы татарской гречихи, мы разработали систему штрих-кодов, используя 50 индексов, выбранных из 26 геномов местных сортов, с использованием стратегии повторного секвенирования. Система двумерных штрих-кодов (рис. 5 и 6) — мощный инструмент для сравнения генотипов образцов татарской гречихи [38–40]. Геномы местных сортов, собранные из шести стран, содержат огромное генетическое разнообразие, адаптированное к широким спектрам условий окружающей среды (Таблица 2). Применяя 50 InDels к этой коллекции, мы обнаружили, что как географические, так и экологические факторы сформировали генетическое разнообразие староместных сортов татарской гречихи (рис. 7).В 73 образцах анализ структуры популяции с использованием данных штрих-кода ДНК выявил их географическое распределение (рис. 8), которое приспособлено к адаптации в разнообразных топографических условиях агроэкосистемы [71–74]. В филогенетическом анализе с использованием несинонимичных SNP 26 образцов показали тесную связь с образцами из Бутана и Пакистана. (Рис 7). Это наблюдение согласуется с предыдущим исследованием, в котором было обнаружено, что анализ генетической структуры популяции с использованием данных SSR сгруппировал одну группу, которая в основном была распространена в Непале, Бутане и регионах плато Юньнань-Гуйчжоу в Китае, а другая группа произошла в основном с Лессового плато. регионы Хунань и Хубэй Китая и США [9].Эти результаты согласуются с ранее предложенным методом обмена семенами на близком географическом расстоянии [5,8]. Вместе с SSR [9], AFLP [41], RAPD [42,43] и ISSR [44] система штрих-кодов с использованием InDels может дополнить текущие исследования и расширить наше понимание как филогенетических сигналов, так и вариаций на уровне популяции [38– 40]. Эти филогенетические и структурные анализы образцов винной гречихи послужат важным ресурсом для более глубокого понимания эволюции клон-специфических генных сетей, связанных со специфическими для окружающей среды особенностями развития и адаптации староместных сортов винной гречихи.
В соответствии с их положением в унигенах 50 InDel были разделены на пять типов: кодирующая область, 5′-нетранслируемая область, 3′-нетранслируемая область, область сплайсинга и межгенная область (таблица S7). InDels может быть склонен к изменениям в экспрессии генов посредством цис-регуляторных мутаций [75]. Постулируется, что InDels некоторых генов, например, RPP4 [76], UGT73B5 [77], TPS4 [78] и DREB3 [79], играют роль в различных ответах на стрессы окружающей среды во время рост и выживание.С доказательствами, связывающими 50 InDels с экологической адаптивностью, эти результаты согласуются с разделением татарской гречихи на различные группы в соответствии с их географическим распределением.
Несмотря на самоопыление винной гречихи через 3 дополнительных поколения с использованием метода SSD, в геномной ДНК 26 потомков было обнаружено около 10% гетерогенных InDel (таблица S3). Предыдущие исследования показали, что фермеры выращивали несколько староместных сортов татарской гречихи, подходящих для различных местных микроклиматов и условий окружающей среды [5], что могло стимулировать поток генов староместных сортов в пределах региона [80].В этом исследовании мы обнаружили сильную поддержку аллогамии среди местных сортов, которая способствует гетерогенным InDels у винной гречихи, что может определять генетическое разнообразие местных сортов в регионе. Учитывая влияние гетерозиготного InDel на увеличение частоты однонуклеотидных мутаций [75,81], это может быть важной характеристикой, которая позволяет татарской гречке адаптироваться к разнообразным топографическим условиям агроэкосистемы. Эти результаты предполагают, что дивергентная экспрессия генов локусов InDel может влиять на его адаптацию к условиям окружающей среды.
Мы обнаружили, что образцы татарской гречихи можно различить по 50 InDels на основе генотипа в зависимости от местоположения с различающейся средой (рис. 7 и 8). Гречиха татарская — это разнообразная культура, у которой в каждом генофонде произошли отдельные события одомашнивания, за которыми последовала расовая и рыночная диверсификация, что привело к различным морфологическим характеристикам в каждом коммерческом рыночном классе [82]. Объединение нескольких типов данных может компенсировать недостающую или ненадежную информацию в одном типе данных.Важно отметить, что полное управление ресурсами может быть обнаружено только в том случае, если при анализе будут учтены генотип, фенотип и хемотип образцов татарской гречихи. Чтобы создать удобную для пользователя общую платформу для ресурсов генотипа, фенотипа и хемотипа, мы планируем объединить данные генотипа с данными этого фенотипа и хемотипа (содержание рутина) для образцов гречихи винного сорта (S3 рис.). Наша платформа может быть использована в генетических исследованиях, селекционных программах и для эффективных систем управления ресурсами гречихи.Повышение урожайности винной гречки потребует адаптации урожая к сельскохозяйственной среде новых мест. Система штрих-кодов может помочь селекционерам винной гречки идентифицировать образцы зародышевой плазмы и может использоваться в качестве родительских доноров для селекции винных сортов гречихи.
Благодарности
Искренне благодарим Мисук Ли за выращивание ресурсов татарской гречки, Джуён Шим за выделение образцов ДНК и Хёнджин Шин за техническую помощь в лаборатории.Авторы хотели бы поблагодарить Enago (www.enago.co.kr) за обзор на английском языке.
Ссылки
- 1. Охниши О., Мацуока Ю. Поиск дикого предка гречихи II. Таксономия видов Fagopyrum (Polygonaceae) на основе морфологии, изоферментов и изменчивости хпДНК. Genes Genet Syst. 1996. 71: 383–390.
- 2. Yasui Y, Ohnishi O. Филогенетические отношения между видами Fagopyrum , выявленные нуклеотидными последовательностями ITS-области гена ядерной рРНК.Genes Genet Syst. 1998. 73 (4): 201–210. pmid: 9880918
- 3. Eggum BO, Kreft I, Javornik B. Химический состав и качество белка гречихи ( Fagopyrum esculentum Moench). Растительная пища Hum Nutr. 1980. 30 (3): 175–179.
- 4. Бонафачча Г., Мароккини М., Крефт И. Состав и технологические свойства муки и отрубей из гречневой крупы обыкновенной и винной. Food Chem. 2003; 80: 9–15.
- 5. Хуанг В., Джарвис Д., Ахмед С., Лонг К.Генетическое разнообразие татарской гречки в Гималаях связано с разнообразием местных фермерских сортов и низкой диетической зависимостью. Устойчивость. 2017; 9 (10): 1806.
- 6. Мурай М., Охниши О. Популяционная генетика культурной гречки обыкновенной, Fagopyrum esculentum Moench. X. Маршруты диффузии, выявленные маркерами RAPD. Genes Genet Syst. 1996. 71: 211–218. pmid: 8979373
- 7. Охниши О. Поиски дикого предка гречихи III. Дикий предок культурной гречки обыкновенной и татарской гречихи.Econ Bot. 1998. 52 (2): 123–133. Доступно на http://www.jstor.com/stable/4256049.pdf. По состоянию на 18 сентября 2020 г.
- 8. Delêtrea M, McKey DB, Hodkinson TR. Брачные обмены, обмены семенами и динамика маниокального разнообразия. Proc Natl Acad Sci U S. A. 2011; 108 (45): 18249–18254. pmid: 22042843
- 9. Хоу С., Сунь З., Линху Б., Сюй Д., Ву Б., Чжан Б. и др. Генетическое разнообразие сортов гречихи ( Fagopyrum tartaricum Gaertn.) Оценивалось с помощью маркеров SSR, разработанных на основе последовательностей геномных исследований.Отчет Plant Mol Biol. 2016; 34 (1): 233–241.
- 10. Kreft I, Fabjani N, Germ M. Rutin в защите растений от гречихи и ее значении для производства функционального питания. Фагопирум. 2003; 20: 7–11. Доступно на https://www.researchgate.net/publication/255602730. По состоянию на 18 сентября 2020 г.
- 11. Бишт И.С., Мехта П.С., Бхандари округ Колумбия. Традиционное разнообразие сельскохозяйственных культур и его сохранение на фермах для устойчивого сельскохозяйственного производства в Кумаон Гималаи штата Уттаранчал: тематическое исследование.Genet Resour Crop Evol. 2007. 54: 345–357.
- 12. Лю М., Ма З., Чжэн Т., Сунь В., Чжан Ю., Цзинь В. и др. Понимание взаимосвязи между физиологическими изменениями и развитием семян татарской гречихи ( Fagopyrum tataricum Gaertn.). BMC Genomics. 2018; 19 (1): 648. pmid: 30170551
- 13. Шпенглер Р. Антропогенное распространение семян: переосмысление истоков одомашнивания растений. Trends Plant Sci. 2020; 25 (4): 340–348. pmid: 32191870
- 14.Беван М.В., Уауи С., Вульф ББХ, Чжоу Дж., Красилева К., Кларк М.Д. Геномные инновации для улучшения сельскохозяйственных культур. Природа. 2017; 543: 346–354. pmid: 28300107
- 15. Ли СК, Чжан Кью. Успехи в разработке функциональных продуктов из гречки. Crit Rev Food Sci Nutr. 2001. 41 (6): 451–464. pmid: 11592684
- 16. Джонс М.К., Лю X. Истоки сельского хозяйства в Восточной Азии. Наука. 2009. 324 (5928): 730–731. pmid: 19423806
- 17. Стедман К.Дж., Бургун М.С., Льюис Б.А., Эдвардсон С.Е., Обендорф Р.Л.Минералы, фитиновая кислота, дубильные вещества и рутин во фракциях помола семян гречихи. J Sci Food Agric. 2001. 81 (11): 1094–1100.
- 18. Fabjan N, Rode J, Košir IJ, Wang Z, Zhang Z, Kreft I. Татарская гречка ( Fagopyrum tataricum Gaertn.) Как источник диетического рутина и кверцитрина. J. Agric Food Chem. 2003. 51 (22): 6452–6455. pmid: 14558761
- 19. Крефт И., Фабьян Н., Ясумото К. Содержание рутина в пищевых материалах и продуктах гречки ( Fagopyrum esculentum Moench).Food Chem. 2006. 98 (3): 508–512.
- 20. Гупта Н., Шарма С.К., Рана Дж.С., Чаухан Р.С. Экспрессия генов биосинтеза флавоноидов через вариацию содержания рутина на разных стадиях роста видов Fagopyrum . J. Plant Physiol. 2011. 168 (17): 2117–2123. pmid: 21872967
- 21. Ван Л, Ян Х, Цинь П, Шан Ф, Рен Г. Флавоноидный состав, антибактериальные и антиоксидантные свойства экстракта винных отрубей гречихи. Ind Crops Prod. 2013; 49: 312–317.
- 22.Hu Y, Hou Z, Liu D, Yang X. Флавоноиды татарской гречки защищают печеночные клетки от окислительного стресса, вызванного высоким уровнем глюкозы, и резистентности к инсулину через сигнальные пути MAPK. Food Funct. 2016; 7 (3): 1523–1536. pmid: 26899161
- 23. Kim SJ, Sohn HB, Kim GH, Lee YY, Hong SY, Kim GD и др. Сравнение и проверка содержания рутина и кверцетина в соответствии с методом экстракции винной гречки ( Fagopyrum tataricum Gaertn.). Корейский J Food Sci Technol.2017; 49 (3): 258–264.
- 24. Джоши Б.К. Корреляционный, регрессионный анализ и анализ путевых коэффициентов для некоторых компонентов урожайности обычной и татарской гречихи в Непале. Фагопирум. 2005. 22: 77–82.
- 25. Ван Y, Кэмпбелл CG. Селекция татарской гречихи ( Fagopyrum tataricum L. Gaertn.) Путем гибридизации с ее рисово-винным типом. Euphytica. 2007. 156 (3): 399–405.
- 26. Судзуки Т., Моршита Т., Мукаса Ю., Такигава С., Йокота С., Исигуро К. и др.Выведение «Мантен-Кирари», не горького и содержащего следы рутинозидазы сорта татарской гречихи ( Fagopyrum tataricum Gaertn.). Breed Sci. 2014. 64 (4): 344–350. pmid: 25914589
- 27. Ли Ц, Се З, Ван И, Лу В, Инь Г, Сунь Д. и др. Корреляционный и генетический анализ толщины оболочки семян и факторов урожайности у гречихи татарской ( Fagopyrum tataricum (L.) Gaertn.). Breed Sci. 2019; 69: 464–470. pmid: 31598079
- 28. Singh SP. Расширение генетической базы культурных сортов фасоли.Crop Sci. 2001; 41: 1659–1675.
- 29. Hyten DL, Song Q, Zhu Y, Choi I.Y, Nelson RL, Costa JM, et al. Влияние генетических узких мест на разнообразие генома сои. Proc Natl Acad Sci U S. A. 2006; 103 (45): 16666–16671. pmid: 17068128
- 30. Бхандари Х. Р., Бхану А. Н., Шривастава К., Сингх М. Н., Шрейя, Хемантаранджан А. Оценка генетического разнообразия сельскохозяйственных культур — обзор. Adv Plants Agric Res. 2017; 7 (3): 279–286.
- 31. Эриксен П.Дж. Концептуализация продовольственных систем для исследования глобального изменения окружающей среды.Glob Environ Change. 2008. 18 (1): 234–245.
- 32. Hughes AR, Inouye BD, Johnson MTJ, Underwood N, Vellend M. Экологические последствия генетического разнообразия. Ecol Lett. 2008. 11 (6): 609–623. pmid: 18400018
- 33. Бриттен Р.Дж., Роуэн Л., Уильямс Дж., Кэмерон Р.А. Большинство расхождений между близкородственными образцами ДНК связано с инделениями. Proc Natl Acad Sci U S. A. 2003; 100 (8): 4661–4665. pmid: 12672966
- 34. Väli Ü, Brandström M, Johansson M, Ellegren H.Инсерционно-делеционные полиморфизмы (индели) как генетические маркеры в природных популяциях. BMC Genet. 2008; 9 (1): 8. pmid: 18211670
- 35. Хоу X, Ли Л., Пэн З., Вэй Б., Тан С., Дин М. и др. Платформа высокоплотных маркеров INDEL / CAPS для картографического клонирования Arabidopsis. Плант Дж. 2010; 63 (5): 880–888. pmid: 20561258
- 36. Могхаддам С.М., Сонг К., Мамиди С., Шмутц Дж., Ли Р., Креган П. и др. Разработка маркеров InDel, специфичных для рыночного класса, на основе данных последовательностей следующего поколения в Phaseolus vulgaris L.Фронтальный завод им. 2014; 5: 185. pmid: 24860578
- 37. Meng HT, Zhang YD, Shen CM, Yuan GL, Yang CH, Jin R и др. Анализ генетического полиморфизма 30 индейцев в китайской этнической группе Xibe и его популяционно-генетических различий с другими группами. Научные отчеты. 2015; 5 (5): 8260. pmid: 25651970
- 38. Sohn HB, Kim SJ, Hwang TY, Park HM, Lee YY, Markkandan K и др. Система штрих-кодов для генетической идентификации сортов сои [ Glycine max (L.) Merrill] с использованием маркеров InDel, специфичных для блоков плотных вариаций.Фронтальный завод им. 2017; 8: 520. pmid: 28443113
- 39. Sohn HB, Song YH, Kim SJ, Hong SY, Kim KD, Koo BC и др. Идентификация и паттерны хромосомных перестановок сортов сои, выращенных в Квангвон-до, с использованием 202 маркеров InDel, специфичных для блоков вариаций. Корейский J Breed Sci. 2018; 50 (4): 396–405.
- 40. Chun JB, Jin M, Jeong N, Cho C, Seo MS, Choi MS и др. Генетическая идентификация и филогенетический анализ новых сортов и 149 корейских сортов с использованием 27 маркеров InDel, выбранных из плотных блоков вариаций сои ( Glycine max (L.) Меррилл). Korean J Plant Res. 2019; 32 (5): 519–542.
- 41. Цуджи К., Охниши О. Филогенетические взаимоотношения между популяциями дикой и культивируемой татарской гречихи ( Fagopyrum tataricum Gaertn.), Выявленные с помощью анализа AFLP. Genes Genet Syst. 2001. 76 (1): 47–52. pmid: 11376551
- 42. Цуджи К., Охниши О. Происхождение культурной татарской гречихи ( Fagopyrum tataricum Gaertn.), Выявленное с помощью RAPD-анализа. Genet Resour Crop Evol. 2000; 47: 431–438.
- 43. Цуджи К., Охниши О. Филогенетическое положение восточно-тибетских природных популяций татарской гречихи ( Fagopyrum tataricum Gaertn.), Выявленное с помощью RAPD-анализа. Genet Resour Crop Evol. 2001. 48 (1): 63–67.
- 44. Чжан А., Чжао Л. Молекулярная характеристика генетического разнообразия малоиспользуемых культур; Гречка как пример. Acta Hortic. 2013; 979: 407–419.
- 45. Кацу К., Сузуки Т., Фуджино К., Моришита Т., Нода Т. Разработка ДНК-маркера для распознавания разновидностей, специфичного для «Manten-Kirari», на основе последовательности NGS-РНК в татарской гречке ( Fagopyrum tataricum ).Food Chem. 2019; 295: 51–57. pmid: 31174789
- 46. Чжан Л., Ма М., Лю Л. Идентификация генетического локуса, лежащего в основе легкого шелушения в рисовом тартаре, для легкой послеуборочной обработки винной гречки. Гены. 2020; 11 (4): 459. pmid: 32340240
- 47. Ясуи Ю., Хиракава Х., Уэно М., Мацуи К., Танака Т.К., Янг С.Дж. и др. Сборка чернового генома гречихи и его применение в идентификации агрономически полезных генов. ДНК Res. 2016; 23 (3): 215–224. pmid: 27037832
- 48.Zhang L, Li X, Ma B, Gao Q, Du H, Han Y и др. Геном татарской гречихи дает представление о биосинтезе рутина и устойчивости к абиотическим стрессам. Завод Мол. 2017; 10 (9): 1224–1237. pmid: 28866080
- 49. Bansal V, Boucher C. Технологии секвенирования и анализ: где мы были и куда мы идем? iScience. 2019; 18: 37–41. pmid: 31472161
- 50. Джанван М., Сревонгчай Т., Срипичитт П. Селекция риса для получения высокой урожайности с помощью усовершенствованного метода селекции методом спуска одного семени.J Plant Sci. 2013; 8 (1): 24–30.
- 51. Halbrecq B, Romedenne P, Ledent JF. Эволюция цветения, созревания и завязывания семян гречихи ( Fagopyrum esculentum Moench): количественный анализ. Europ J Agronomy. 2005; 23: 209–224.
- 52. Рейес-Вальдес М.Х., Сантакрус-Варела А., Мартинес О., Симпсон Дж., Хаяно-Канаширо, С., Кортес-Ромеро С. Анализ и оптимизация общего отбора образцов ДНК с бинарной оценкой для характеристики зародышевой плазмы. PLoS ONE. 2013; 8: e79936.pmid: 24260321
- 53. Роджерс С.О., Бендич А.Дж. Извлечение общей клеточной ДНК из растений, водорослей и грибов. В: Гельвин С.Б., Шильпероорт Р.А., редакторы. Руководство по молекулярной биологии растений. Спрингер: Дордрехт; 1994. стр. 183–190. https://doi.org/10.1007/978-94-011-0511-8_12
- 54. Chin CS, Peluso P, Sedlazeck FJ, Nattestad M, Concepcion GT, Clum A и др. Поэтапная диплоидная сборка генома с секвенированием одной молекулы в реальном времени. Нат методы. 2016; 13: 1050–1054. pmid: 27749838
- 55.Луо Р., Лю Б., Се И, Ли З, Хуанг В., Юань Дж. И др. SOAPdenovo2; Эмпирически улучшенный ассемблер de novo с эффективным использованием памяти. Gigascience. 2012; 1 (1): 18. pmid: 23587118
- 56. Хуанг С., Кан М., Сюй А. Гапломергеры: восстановление обеих гаплоидных подгрупп из сборки диплоидного генома с высокой гетерозиготностью. Биоинформатика. 2017; 33 (16): 2577–2579. pmid: 28407147
- 57. Бетцер М., Хенкель К.В., Янсен Х.Дж., Батлер Д., Пировано В. Строим предварительно собранные контиги лесами с использованием SSPACE.Биоинформатика. 2011. 27 (4): 578–579. pmid: 21149342
- 58. Ли Х, Дурбин Р. Быстрое и точное согласование короткого чтения с преобразованием Барроуза-Уиллера. Биоинформатика. 2009. 25 (14): 1754–1760. pmid: 19451168
- 59. Маккенна А., Ханна М., Бэнкс Э., Сиваченко А., Цибульскис К., Керницкий А. и др. Набор инструментов для анализа генома: платформа MapReduce для анализа данных секвенирования ДНК следующего поколения. Genome Res. 2010. 20 (9): 1297–1303. pmid: 20644199
- 60. Андерсон Дж. А., Черчилль Г. А., Аутриг Дж. Э., Танксли С. Д., Сорреллс Мэн.Оптимизация родительского отбора для карт генетического сцепления. Геном. 1993. 36 (1): 181–186. pmid: 18469981
- 61. Лю К., Муза С.В. PowerMarker: интегрированная среда анализа для анализа генетических маркеров. Биоинформатика. 2005. 21 (9): 2128–2129. pmid: 15705655
- 62. Perrier X, Jacquemoud-Collet JP. Программное обеспечение DARwin. 2006. Доступно на http://darwin.cirad.fr/.
- 63. Причард Дж. К., Стивенс М., Розенберг Н. А., Доннелли П. Картирование ассоциаций в структурированных популяциях.Am J Hum Genet. 2000. 67 (1): 170–181. pmid: 10827107
- 64. Kim YH, Park HM, Hwang TY, Lee SK, Choi MS, Jho S и др. Метод вариационной блочной геномики сельскохозяйственных культур. BMC Genomics 2014; 15: 477. pmid: 24929792
- 65. Цзян П., Бурчински Ф., Кэмпбелл С., Пирс Г., Австрия Дж. А., Бриггс С.Дж. Содержание рутина и флавоноидов у трех видов гречихи Fagopyrum esculentum , F . tataricum и F . homotropicum и их защитное действие от перекисного окисления.Food Res Int. 2007. 40 (3): 356–364.
- 66. Крефт С., Струкель Б., Габерчик А., Крефт И. Рутин в травах гречихи, выращенных при разных уровнях УФ-В излучения: сравнение двух УФ-спектрофотометрических данных и метода ВЭЖХ. J Exp Bot. 2002. 53 (375): 1801–1804. pmid: 12147730
- 67. Мацуура Х., Инанага С., Мурата К. Различия в вегетативном росте между обыкновенной и татарской гречкой в солевой гидропонной культуре. Plant Prod Sci. 2005. 8 (5): 533–538.
- 68.Оуэнс Д.К., Алердинг А.Б., Кросби К.С., Бандара А.Б., Вествуд Д.Х., Винкель Б.С. Функциональный анализ предсказанного семейства генов флавонолсинтазы у Arabidopsis. Plant Physiol. 2008; 147: 1046–1061. pmid: 18467451
- 69. Germ M, Brenznik B, Dolinar N, Kreft I., Gaberščik A. Комбинированный эффект ограничения воды и УФ-B излучения на гречневую крупу обыкновенную и винную. Cereal Res Commun. 2013. 41 (1): 97–105.
- 70. Пандей В., Ниранджан А., Атри Н., Чандрашекхар К., Мишра М.К., Триведи П.К., Мисра П.Ген WsSGTL1 из Withania somnifera , модулирует профиль гликозилирования, антиоксидантную систему и придает трансгенному табаку устойчивость к биотическому и солевому стрессу. Planta. 2014. 239 (6): 1217–1231. pmid: 24610300
- 71. Джексон Л., ван Нордвейк М., Бентссон Дж., Фостер В., Липпер Л., Пуллеман М. и др. Биоразнообразие и устойчивость сельского хозяйства: от оценки к адаптивному управлению. Curr Opin Environ Sustain. 2010; 2: 80–87.
- 72. Вигуру Y, Барно А., Скарчелли Н., Тюйе А.Биоразнообразие, эволюция и адаптация культурных растений. C R Biol. 2011; 334: 450–457. pmid: 21640954
- 73. Ахмед С., Петерс С.М., Лонг К.Л., Мейер Р., Уначукву У., Литт А. и др. Биоразнообразие и фитохимическое качество в местных и поддерживаемых государством системах управления чаем в Юньнани, Китай. Conserv Lett. 2013; 6: 28–36.
- 74. Цюй Й, Чжоу Й, Ван З, Ван З, Ван П, Гао Дж и др. Анализ генетического разнообразия и структуры ресурсов татарской гречихи из регионов-производителей.Scientia Agricultura Sinica. 2016; 49 (11): 2049–2062.
- 75. Тиан Д., Ван Ц., Чжан П., Араки Х., Ян С., Крейтман М. и др. Частота однонуклеотидных мутаций увеличивается ближе к вставкам / делециям у эукариот. Природа. 2008. 455: 105–108. pmid: 18641631
- 76. ван дер Бизен Э.А., Фредди К. Т., Кан К., Паркер Дж. Э., Джонс Д. Д. Г.. Arabidopsis RPP4 является членом мультигенного семейства RPP5 генов TIR-NB-LRR и придает устойчивость к ложной мучнистой росе посредством множества компонентов передачи сигналов.Плант Дж. 2002; 29 (4): 439–451. pmid: 11846877
- 77. Langlois-Meurinne M, Gachon CM, Saindrenan P. Чувствительная к патогену экспрессия генов гликозилтрансфераз UGT73B3 и UGT73B5 необходима для устойчивости к Psudomonas syringae pv томат в Arabidopsis. Plant Physiol. 2005. 139 (4): 1890–1901. pmid: 16306146
- 78. Attaran E, Rostás M, Zeier J. Pseudomonas syringae вызывает выброс терпеноида (E, E) -4,8,12-триметнил-1,3,7,11-тридекатетраена в листьях Arabidopsis посредством передачи сигналов жасмонатом и экспрессия терпен-синтазы TPS4.Мол Растительный Микроб Взаимодействовать. 2008. 21 (11): 1482–1497. pmid: 18842097
- 79. Ислам MS, Ван MH. Экспрессия белка-3, связывающего элемент, реагирующий на дегидратацию ( DREB3 ) при различных абиотических стрессах в томате. BMB Rep. 2009; 42 (9): 611–616. pmid: 19788864
- 80. Беллуччи Э, Биточчи Э, Рау Д., Нанни Л., Феррадини Н., Джардини А. и др. Структура популяций староместных сортов ячменя и генетический поток современных сортов. PLoS ONE. 2013; 8 (12): e83891.pmid: 24386303
- 81. Холлистер Дж. Д., Росс-Ибара Дж., Гаут Б. С.. Частота мутаций, связанных с Indel, зависит от системы спаривания у цветковых растений. Mol Biol Evol. 2010. 27 (2): 409–416. pmid: 19825943
- 82. Лейшман М.Р., Вестоби М., Хурадо Э. Корреляты изменения размера семян: сравнение пяти флор умеренного пояса. J Ecol. 1995. 83 (3): 517–529.
Сравнительный клеточный, физиологический и транскриптомный анализы выявили потенциально легкий механизм шелушения рисовой гречневой крупы (Fagopyrum Tararicum) | BMC Plant Biology
Фенотипические и физиологические изменения
По сравнению с обычным сортом татарской гречихи «JQ» созревшая лузга рисово-татарской гречихи «XMQ» показала заметно пониженную механическую прочность, которая легко ломалась и осыпалась при трении рукой (Рис. .1а). Сканирующая электронная микроскопия показала, что на корпусе «XMQ» появилось много дефектов, которых не было на «JQ» (рис. 1b, c). Эти результаты показали, что более низкая механическая прочность корпуса «XMQ» способствовала его легкому шелушению, а признак легкости шелушения был переопределен как признак «хрупкий корпус». Для дальнейшего изучения того, была ли более низкая механическая прочность корпуса «XMQ» вызвана изменением состава корпуса, мы измерили содержание целлюлозы, гемицеллюлозы и лигнина в корпусе «XMQ», «JQ» и других трех обычных сортов татарской гречихи. («JJQ», «PQ» и «CQ»).Как показано на рис. 2, по сравнению с четырьмя нормальными сортами винной гречки, содержание целлюлозы в шелухе «XMQ» было значительно ниже, чем во всех протестированных обычных сортах винной гречихи, в то время как содержание гемицеллюлозы и лигнина в шелухе в «XMQ» было значительно ниже. ниже, чем у двух обычных винных сортов гречихи. Эти результаты свидетельствуют о том, что пониженное содержание целлюлозы, гемицеллюлозы и лигнина может способствовать хрупкости и легкости шелушения рисовой и винной гречихи, основной причиной чего может быть пониженное содержание целлюлозы.
Рис. 1Фенотипы зрелой оболочки семян XMQ и JQ. a Фенотипы зрелых семян XMQ и JQ после ручного лущения 10 раз. b Сканирующие электронные микрофотографии зрелых семян XMQ и JQ. c Сканирующие электронные микрофотографии зрелой оболочки семян XMQ и JQ
Рис. 2Содержание целлюлозы, гемицеллюлозы и лигнина в оболочке семян различных сортов винной гречихи. Значения представляли собой средние значения трех независимых биологических повторов, а столбцы с разными буквами представляли значимые различия ( P <0.05)
Динамическое изменение содержания целлюлозы и гемицеллюлозы и толщины корпуса в «XMQ» и «JQ»
Для исследования накопления целлюлозы и гемицеллюлозы во время развития корпуса, содержания целлюлозы и гемицеллюлозы в оболочке семян «XMQ» и «JQ» определялись на четырех различных этапах (рис. 3а). Как показано на рис. 3b, c, два сорта продемонстрировали сходную тенденцию в изменении содержания целлюлозы и гемицеллюлозы, которые устойчиво росли во время развития корпуса.Кроме того, значительная разница в содержании целлюлозы и гемицеллюлозы между двумя сортами начала наблюдаться при 10 DAP. Для дальнейшего понимания того, изменяется ли толщина канавы корпуса у двух сортов во время развития семян, был проведен анализ парафинового сечения, а толщина канавы корпуса была измерена в соответствии с результатами парафинового сечения (рис. 4). Толщина корпусной канавки «XMQ» значительно уменьшилась во время развития семян, в то время как толщина корпусной канавы «JQ» уменьшилась только на 10 DAP по сравнению с 5 DAP, и не наблюдалось очевидной разницы после 10 DAP (Рис.4б). Кроме того, толщина канавы корпуса «XMQ» была значительно ниже, чем у «JQ» на 15 и 20 DAP, хотя толщина его корпуса была явно выше, чем у «JQ» на 5 и 10 DAP (рис. 4b). . Просвечивающая электронная микроскопия также показала, что толщина клеточной стенки клеток склеренхимы корпуса «XMQ» была значительно меньше, чем у «JQ» (рис. 5). Эти результаты показали, что содержание целлюлозы, гемицеллюлозы и толщина канавки корпуса «XMQ» и «JQ» были динамическими и разными изменениями, и эти образцы корпуса можно было использовать для дальнейших анализов транскриптома.
Рис. 3Фенотипы семян ( a ), целлюлоза оболочки ( b ) и содержание гемицеллюлозы ( c ) в XMQ и JQ. Звездочки указывают на значительные различия (** P <0,01)
Рис. 4Микрофотографии сечения парафина семян ( a ) и толщины канавы корпуса ( b ) XMQ и JQ. Столбики с разными буквами представляли значимые различия ( P <0,05) у одной и той же разновидности на разных стадиях развития.Звездочки указывают на значительные различия (** P <0,01) между двумя разновидностями на одной и той же стадии развития
Рис. 5Анатомические особенности клеток склеренхимы в оболочке семян 15 DAP JQ и XMQ. a, и c, — микрофотографии клеточных стенок склеренхимы в трансмиссии JQ. b и d электронные микрофотографии стенок клеток склеренхимы в XMQ. e — толщина клеточной стенки клеток склеренхимы в JQ и XMQ.Звездочки указывают на существенные различия (** P <0,01) между JQ и XMQ
Транскриптомный анализ развития шелухи у сортов татарской гречихи
Чтобы понять динамику транскриптома во время развития корпуса, RNA-Seq анализирует корпус «XMQ». »И« JQ »были выполнены на четырех различных стадиях разработки (8 тканей, три независимых биологических повтора для каждой ткани, всего 24 образца). Всего 521,39 и 517,72 миллиона чистых чтений, в среднем 43.43 и 43,09 миллиона чистых чтений для каждого образца были сгенерированы для «XMQ» и «JQ» соответственно (дополнительный файл 1: таблица S1). От 85,84 до 92,74% чистых считываний были сопоставлены с эталонным геномом для каждого образца, из которых 64,54–73,47% были картированы однозначно. Всего 29 843 генных локуса, включая 28 465 известных и 1378 новых генных локусов, были сгенерированы из всех картированных чистых считываний 24 образцов с помощью анализов Cufflinks и Cuffmerge. PCC между биологическими репликами разных тканей изменилось с 0.От 90 до 0,99 (за исключением одной реплики J15 и J20 в «JQ» и одной реплики X15 в «XMQ», которые были отфильтрованы для дальнейшего анализа), что указывает на высокую надежность и воспроизводимость реплик (дополнительный файл 2: Рисунок S1 ).
Рассчитывали нормализованный уровень экспрессии (FPKM) для каждого гена, и гены с FPKM ≥0,1 считали экспрессированными. В общей сложности 27 955 генов, включая 26 589 известных и 1366 новых генов, были экспрессированы по крайней мере в одном образце. Среди них количество экспрессируемых генов в различных тканях изменилось с 23 575 (X20) до 25014 (X10) в «XMQ» и от 23 282 (J20) до 24 266 (J5) в «JQ» (дополнительный файл 2: рисунок S2a).Доля генов, представляющих очень высокую (FPKM ≥100), высокую (50 ≤ FPKM <100), умеренную (10 ≤ FPKM <50), низкую (2 ≤ FPKM <10) и очень низкую (0,1 ≤ FPKM <2) экспрессия была относительно сходной во всех тканях, за исключением ткани X20 (дополнительный файл 2: рисунок S2b). Кроме того, количество генов с очень низкой экспрессией (около 42%) и с умеренной экспрессией (около 30%) составляло две самые большие доли во всех тканях (дополнительный файл 2: рисунок S2b). Взятые вместе, эти анализы показали, что мы получили достаточный охват транскриптома оболочки развития у этих двух сортов татарской гречихи.
Сравнение транскриптомов развития корпуса у сортов винной гречихи выявило критические стадии различий в развитии корпуса.
Для исследования взаимосвязей между транскриптомами развития корпуса из «XMQ» и «JQ», HCA (дополнительный файл 2: Рисунок S3a) и PCA (дополнительный файл 2: рисунок S3b) были выполнены на основе средних значений FPKM для 27 955 генов, которые были экспрессированы по крайней мере в одном из восьми образцов ткани. В результате наблюдалась более высокая корреляция одной и той же стадии развития между двумя сортами, за исключением 20 DAP (Рисунок S3a).Анализ PCA показал, что X5 и J5 были сгруппированы вместе, в то время как между X10 и J10, X15 и J15, а также X20 и J20 наблюдалось четкое разделение (дополнительный файл 2: рисунок S3b), что свидетельствует о более высокой степени сходства транскрипционных программ между этими двумя сорта на 5 DAP и очевидные транскрипционные различия существовали на 10, 15 и 20 DAP. Кроме того, эти анализы также показали, что 10 DAP может быть критическим этапом для определения образования различий в оболочке между двумя сортами на молекулярном уровне.
Идентификация генов, которые специфически / предпочтительно экспрессируются на каждой стадии развития корпуса у обоих сортов
Гены, которые специфически / предпочтительно экспрессируются на каждой стадии развития корпуса у обоих сортов, были идентифицированы на основе алгоритма SS с оценкой SS ≥ 0,5. В общей сложности 6649 и 5918 специфических / предпочтительных генов были идентифицированы на всех четырех стадиях для «JQ» и «XMQ», соответственно. Среди этих генов 348 (5,2%) и 302 (5,1%) специфических / предпочтительных гена кодировались факторами транскрипции (TF).Количество специфичных для стадии / предпочтительных генов варьировало от 392 до 4613 для «JQ» и от 267 до 3959 для «XMQ» (рис. 6а). 5 DAP и 10 DAP имели наибольшее и наименьшее количество специфичных для стадии / предпочтительных генов, соответственно, для двух сортов (рис. 6a). Кроме того, высокая доля специфичных для стадии / предпочтительных генов была общей для обоих сортов, а гены, специфичные для сорта, также наблюдались на всех четырех стадиях двух сортов (рис. 6а). Тепловая карта всех этих стадийно-специфичных / преимущественно экспрессируемых генов у двух сортов татарской гречихи показана на рис.6б. Эти результаты показали, что каждая стадия развития корпуса имела свои собственные независимые и общие программы развития для обоих сортов, а также точно отражала накопление эндогенных мРНК в развитии корпуса двух сортов.
Рис. 6Предпочтительная / стадийно-специфическая экспрессия генов во время развития корпуса у XMQ и JQ. a Число специфических и общих, предпочтительно экспрессируемых генов в XMQ и JQ на каждой стадии развития. b Тепловая карта экспрессии преимущественно экспрессируемых генов на разных стадиях развития в XMQ и JQ. c Карта обогащения GO (биологический процесс) преимущественно экспрессируемых генов на 10 DAP развития корпуса в XMQ и JQ
Анализ обогащения онтологии генов (GO) был проведен для специфичных для стадии / предпочтительных генов двух сортов при каждый этап. Результаты обогащения биологического процесса показали, что 5 DAP развития корпуса были отмечены клеточным циклом и процессом клеточного биосинтеза (дополнительный файл 2: Рисунок S4), 10 DAP был значительно и специфически отмечен клеточной стенкой и метаболическим процессом фенилпропаноидов (Рис.6c), 15 DAP был главным образом отмечен «метаболизмом серы» (Дополнительный файл 2: Рисунок S5), а основной 20 DAP был вовлечен в метаболизм липидов, ответ на биотический и абиотический стресс и транспорт (Дополнительный файл 2: Рисунок S6). Эти результаты свидетельствуют о том, что эти специфичные для стадии / предпочтительные гены выполняли специфические для стадии функции во время развития шелухи винной гречихи, и 10 DAP был ключевым этапом в биогенезе детерминации клеточной стенки шелухи винной гречихи.
ДГЭ двух сортов гречневой крупы
Гены, экспрессия которых существенно различалась между «XMQ» и «JQ», были идентифицированы на каждой стадии развития шелухи.В общей сложности 9250 (включая 693 гена, кодирующие TF) и 4187 (294 гена, кодирующие TF) показали значительно более высокую и более низкую экспрессию на разных стадиях развития корпуса у «XMQ» по сравнению с «JQ», соответственно (рис. 7) . Между двумя сортами два наибольших числа DEG имели место при 20 DAP (5884) и 10 DAP (3916) (фиг. 7), в то время как наименьшее количество наблюдалось при 15 DAP. Кроме того, некоторые члены большинства семейств TF показали значительно различающееся выражение в «XMQ», а количество различных семейств TF показало очевидное различие, что означает, что они выполняли различные функции во время разработки корпуса (дополнительный файл 2: рисунок S7).
Рис. 7Различные гены экспрессии в XMQ по сравнению с JQ на разных стадиях развития корпуса. Пурпурное число представляет количество TF с повышенной и пониженной регуляцией.
Для дальнейшего исследования биологических функций этих DEG между «XMQ» и «JQ» были выполнены анализы обогащения GO. В общей сложности 159, 250, 92 и 161 биологический процесс были значительно обогащены ( P <0,05) на 5 DAP, 10 DAP, 15 DAP и 20 DAP, соответственно (дополнительный файл 1: Таблица S2).Среди них некоторые биологические процессы были особенно / обычно чрезмерно представлены на разных стадиях развития корпуса (Дополнительный файл 1: Таблица S2). Примечательно, что такие термины GO, как биогенез первичной клеточной стенки растительного типа (GO: 0009833), процесс метаболизма целлюлозы (GO: 0030243), процесс биосинтеза целлюлозы (GO: 0030244) и катаболический процесс целлюлозы (GO: 0030245), были уникально обогащены. при 5 DAP, тогда как метаболический процесс гемицеллюлозы (GO: 0010410) и модификация клеточной стенки (GO: 0042545) были специально обогащены на 10 DAP (дополнительный файл 1: Таблица S2).Кроме того, биогенез клеточной стенки (GO: 0042546) и расхождение плодов (GO: 0010047) были значительно обогащены на 5 DAP и 10 DAP (дополнительный файл 1: таблица S2). Эти результаты показывают, что 5 DAP и 10 DAP были ключевыми стадиями, которые определили разницу в шелухе между двумя сортами татарской гречихи.
Идентификация и оценка ключевых генов, участвующих в различии в скорлупе двух бульваров татарской гречихи, с помощью анализа коэкспрессии генов
Для исследования регуляторной сети генов (GRN) во время развития корпуса и определения ключевых генов, участвующих в различии в корпусе между « XMQ »и« JQ », 9549 генов, которые по-разному экспрессировались по крайней мере на одной стадии развития корпуса между двумя сортами, были использованы для выполнения WGCNA.Всего было идентифицировано 18 модулей (состоящих из 34–3317 генов) и помеченных разными цветами (рис. 8а). Корреляционный анализ этих модулей с содержанием целлюлозы и гемицеллюлозы показал, что модуль MEbrown показал более высокую корреляцию с содержанием целлюлозы (r = 0,8, p = 0,02) (дополнительный файл 2: рисунок S8). В этом модуле гены имели специфически высокую экспрессию в оболочке «JQ» на 20 DAP. На основании гомологичной аннотации в базе данных Arabidopsis TAIR не было обнаружено, что ген в этом модуле участвует в биосинтезе целлюлозы и клеточной стенки, что может быть вызвано видимой разницей в корпусе между двумя культурами, сформировавшимися при 20 DAP.Учитывая, что (1) легкое шелушение является рецессивным признаком, (2) жесткое шелушение является соответствующим доминирующим признаком и (3) 10 DAP было идентифицировано как ключевой этап формирования разницы между двумя сортами, мы предположили, что модуль с гены, которые проявляли специфически высокую экспрессию в «JQ» на 10 DAP, были ключевым модулем, участвующим в формировании различной оболочки двух сортов. В результате было обнаружено, что модуль MEred соответствует этим стандартам и состоит из 533 генов (рис. 8b).На основании гомологичной аннотации в базе данных Arabidopsis TAIR, в этом модуле было идентифицировано 28 TF, которые представляли 17 семейств TF. Все 28 TF были далее идентифицированы как центральные TF из-за высокосвязных узлов в этом модуле. Эти ТФ концентратора включали гомологи ТФ A. thaliana с известными функциями регуляции биосинтеза SCW, такие как регуляторы NAC первого уровня ( NST1 , NST2 и NST3 / SND1 ), второго — регулятор уровня ( MYB46 / MYB83 ) и регуляторы третьего уровня ( MYB54 , MYB103 , C3h24 и C3h25 ) (дополнительный файл 1: Таблица S3).Гомологи XND1 и VNI2 , двух NAC TFs, которые регулируют образование ксилемных сосудов, также были идентифицированы в этих центральных TF (фиг. 9, дополнительный файл 1: таблица S4). Кроме того, было обнаружено, что несколько других узловых ТФ гомологичны ТФ A. thaliana , которые участвуют в передаче сигналов этилена ( EIN3, , ERF71, и RAV ), биосинтезе гиббереллина ( ATh2 ), жасмоническом кислотная передача сигналов ( WRKY50 ) и множественные гормональные реакции ( MIF2 ).Примечательно, что в этом модуле также были идентифицированы 6, 12, 1 и 5 генов ферментов биосинтеза SCW (рис.9, дополнительный файл 1: таблица S3), которые были гомологами известных нижестоящих мишеней ТФ биосинтеза A. thaliana SCW (рис. NAC и MYB) и катализируемый биосинтез целлюлозы, биосинтез гемицеллюлозы, биосинтез пектина и биосинтез лигнина, соответственно. Среди этих генов ферментов четыре гена биосинтеза целлюлозы ( CESA4 , CESA7 , CESA8 и XTh32 ) и восемь генов биосинтеза гемицеллюлозы ( IRX8 , IRX9 , IRX14-L , IRX14-L , IRX14-L , GUX5 , TBL3 , TBL31 и TBL33 ) также были идентифицированы как гены-концентраторы в этом модуле (рис.9). Для сравнения, гены ферментов биосинтеза пектина и лигнина не были идентифицированы как гены-концентраторы в этом модуле (рис. 9).
Рис. 8Сеть коэкспрессии во время развития корпуса в XMQ и JQ. — Иерархическое дерево кластеризации (дендрограмма) всех различных генов экспрессии из XMQ и JQ на основе сетевого анализа коэкспрессии. b Идентифицированный модуль MEred со специфическими генами высокой экспрессии в оболочке JQ на 10 DAP
Рис. 9Сеть коэкспрессии генов из модуля MEred.Желтые, розовые и красные узлы представляют идентифицированные регуляторы первого, второго и третьего уровней биосинтеза SCW, соответственно. Зеленые узлы представляют идентифицированные структурные гены биосинтеза SCW. Узлы павлиньего синего цвета представляют другие идентифицированные TF концентраторов. Синие узлы представляли другие гены. Более крупные узлы указывали на гены-концентраторы, которые имели узлы с высокой степенью связи в этом модуле
Экспрессия наиболее идентифицированных генов-концентраторов TF и генов ферментов биосинтеза SCW была значительно выше в «JQ», чем в «XMQ» при 5 DAP или 10 DAP. или оба 5 DAP и 10 DAP (Дополнительный файл 1: Таблица S4, Дополнительный файл 1: Таблица S5), что согласуется с приведенными выше результатами обогащения GO и снова предполагает, что ранние стадии разработки корпуса были ключевыми в определении разницы в корпусе между двумя сортами гречихи по-татарски.Экспрессия 17 идентифицированных генов, связанных с SCW, включая 7 регуляторных генов и 10 генов ферментов, была дополнительно подтверждена как очень похожая (r ≥ 0,78) на экспрессию, наблюдаемую в данных RNA-seq с помощью анализов RT-qPCR (рис. 10). Это указывало на надежность транскриптомных данных и выявленных генов, обусловивших различие в скорлупе двух сортов татарской гречихи. Для дополнительной проверки того, что различная экспрессия этих идентифицированных генов биосинтеза SCW на ранних стадиях развития шелухи была причиной образования разницы между рисом и винной гречкой и нормальной винной гречкой, экспрессией регуляторов первого слоя ( NST1 , NST2 и SND1 / NST3 ) биосинтеза SCW тестировали в лузге трех других нормальных сортов татарской гречихи.Как показано на рис. 11, все три гена показали наивысшую экспрессию у нормальных сортов винной гречихи при 10 DAP, а значительно различающаяся экспрессия (кратное изменение> 2) между сортами «XMQ» и нормальной винной гречихой наблюдалась как при 5 DAP, так и 10 DAP. Это предполагает, что различная экспрессия этих генов биосинтеза SCW на ранних стадиях развития шелухи была основной причиной различий в шелухе между рисово-винной гречкой и нормальной винной гречкой.
Фиг.10Корреляция между профилями экспрессии идентифицированных генов биосинтеза SCW. Изменение кратности экспрессии генов использовали для построения тепловых карт. J5 использовался в качестве контроля. Левая тепловая карта представляет RNA-seq, а правая тепловая карта представляет RT-qPCR
Рис. 11Экспрессия регуляторных генов первого уровня биосинтеза SCW в шелухах различных сортов винной гречихи
Анализ промотора «In silico» ТФ-концентраторов и ферментных генов, участвующих в биосинтезе SCW, выявили наличие сайтов связывания SCW-связанных TF
Цис-элементы связывания SCW-связанных TF цис-элементов SNBE (связывание NAC) [22] и SMRE (связывание MYB) [23] были исследовали промоторные последовательности девяти SCW-связанных ТФ ( NST1 , NST2 , SND1 / SNT3 , MYB54 , MYB46 / MYB83 , MYB103 (2), C3h24 (2), C3h24 и C3h25 ), четыре гена ферментов, связанных с биосинтезом целлюлозы ( CESA4 , CESA7 , CESA8 и XTh32 ), восемь генов ферментов, связанных с биосинтезом гемицеллюлозы ( IRX8 , IRX9 , IRX14-L , GXM1 , GUX5 , TBL3 , TBL31 и TBL33 ) и другие 19 идентифицированных узловых TF.В результате цис-элементы SNBE и SMRE представлены в промоторных последовательностях почти всех SCW-связанных ТФ и генов ферментов (Таблица 1). Кроме того, большое количество цис-элементов SNBE также было обнаружено в промоторных последовательностях других 19 узловых ТФ, в то время как цис-элементы SMRE не отображались в этих центральных ТФ (таблица 1). Из-за этиленового ответа EIN3 TF был обнаружен в центральных TF, и было продемонстрировано, что он играет решающую регуляторную роль во многих процессах развития [24], поэтому мы дополнительно исследовали цис-элемент, связывающий EIN3, в промоторных последовательностях вышеуказанных генов.Как показано в таблице 1, цис-элемент, связывающий EIN3, появился в промоторных последовательностях 33 из 40 генов, в которых наибольшее количество связывающих EIN3 цис-элементов было обнаружено в регуляторах NAC первого уровня биосинтеза SCW ( NST1 , NST2 и NST3 / SND1 ) (15, 15 и 9) и регулятор формирования ксилемных сосудов XND1 (11), предполагая, что EIN3 может быть прямым регулятором восходящего потока регуляторы NAC первого уровня биосинтеза SCW.
Таблица 1 Количество цис-элементов, связанных с клеточной стенкой (SNBE и SMRE) и связывающих EIN3, представленных в промоторных последовательностях идентифицированных узловых генов в модуле MEredСоциальное и экологическое влияние на татарскую гречиху (Fagopyrum tataricum Gaertn.) сортовое разнообразие в Юньнани, Китай
Альварес Н., Гарин Э., Хасах С., Дуниас Э, Хоссарт-Макки М., Макки Д. (2005) Практика фермеров, динамика метапопуляции и сохранение сельскохозяйственного биоразнообразия на ферме: тематическое исследование сорго среди дуупа в Камеруне к югу от Сахеля.Biol Conserv 121 (4): 533–543
Статья Google Scholar
Баджрачарья Дж., Стил К.А., Джарвис Д.И., Стхапит Б.Р., Уиткомб Дж.Р. (2006) Разнообразие староместных сортов риса в Непале: изменчивость агроморфологических признаков и маркеров SSR у староместных сортов с высокогорного участка. Полевые культуры Res 95 (2–3): 327–335
Статья Google Scholar
Баджрачарья Дж., Браун AHD, Джоши Б.К., Пандай Д., Бания Б.К., Стхапит Б.Р., Джарвис Д.И. (2011) Традиционное семеноводство и генетическое разнообразие сортов ячменя в высокогорных агроэкосистемах Непала.Genet Resour Crop Evol 59 (3): 389–398
Статья Google Scholar
Bellon MR (2004) Концептуализация вмешательств для поддержки сохранения генетических ресурсов на фермах. World Dev 32 (1): 159–172
Статья Google Scholar
Bellon MR, Aguirre A, Berthaud J, Smale M, Dıaz J, Taba S, Aragon FC, Castro H (2003) Совместная селекция местных сортов для сохранения на ферме: пример из Центральных долин Оахаки, Мексика.Genet Resour Crop Evol 50 (4): 401–416
Статья Google Scholar
Campbell C (1997) Гречка. Fagopyrum esculentum Moench. Содействие сохранению и использованию недостаточно используемых и забытых культур 19. Институт генетики растений и исследований сельскохозяйственных культур, Гатерслебен / Международный институт генетических ресурсов растений, Рим
Google Scholar
Кэмпбелл С. (2003) Улучшение посевов гречихи.Фагопирум 20: 1–6
Google Scholar
Chybicki IJ, Oleksa A, Burczyk J (2011) Повышенный инбридинг и сильная структура родства у Taxus baccata , оцененная как по данным AFLP, так и SSR. Наследственность 107 (6): 589–600
CAS Статья PubMed PubMed Central Google Scholar
Earl DA, VonHoldt BM (2012) STRUCTURE HARVESTER: веб-сайт и программа для визуализации выходных данных STRUCTURE и реализации метода Эванно.Conserv Genet Resour 4 (2): 359–361
Статья Google Scholar
Эванно Г., Регнаут С., Гуде Дж. (2005) Определение количества групп людей с помощью программного обеспечения СТРУКТУРА: исследование моделирования. Мол Экол 14 (8): 2611–2620
CAS Статья PubMed Google Scholar
Fofana IJ, Silue S, Diarrassouba N, Kadio AA, Sangare A (2013) Сравнительный анализ полиморфизма длины амплифицированного фрагмента (AFLP) и простого повтора последовательности (SSR) в генетическом разнообразии тика ( Tectona grandis L.е). Int J Adv Agric Res 1: 114–123
Google Scholar
Fogelqvist J, Niittyvuopio A, Ågren J, Savolainen O, Lascoux M (2010) Скрытая генетическая структура популяции: количество предполагаемых кластеров зависит от размера выборки. Mol Ecol Resour 10 (2): 314–323
Статья PubMed Google Scholar
Gill-Langarica H (2011) Анализ генетического разнообразия фасоли обыкновенной на основе молекулярных маркеров.Genet Mol Biol 34 (4): 595–605
CAS Статья PubMed PubMed Central Google Scholar
Хартл Д.Л., Кларк А.Г. (1997) Принципы популяционной генетики. Sinauer Associates, стр. 542
Гласна Цепкова П., Яновска Д., Стехно З. (2009) Оценка генетического разнообразия отобранных образцов винной крупы и гречихи обыкновенной. Span J Agric Res 4: 844–854
Статья Google Scholar
Holland BR, Clarke AC, Meudt HM (2008) Оптимизация параметров автоматической оценки AFLP для улучшения филогенетического разрешения.Syst Biol 57 (3): 347–366
Статья PubMed Google Scholar
Hou Y, Zhang Z, Wu B, Li Y (2009) Анализ генетического разнообразия винной гречихи с использованием маркеров AFLP. Agric Sin 42 (12): 4166–4174
CAS Google Scholar
Якобссон М., Розенберг Н.А. (2007) CLUMPP: программа сопоставления и перестановки кластеров для работы с переключением меток и мультимодальностью в анализе структуры популяции.Биоинформатика 23 (14): 1801–1806
CAS Статья PubMed Google Scholar
Jensen HR, Belqadi L, De Santis P, Sadiki M, Jarvis DI, Schoen DJ (2012) Пример сетей обмена семенами и потока генов для ячменя ( Hordeum vulgare subsp. vulgare ) в Марокко . Genet Resour Crop Evol 60 (3): 1119–1138
Статья Google Scholar
Джомбарт Т., Ахмед I (2011) Adegenet 1.3-1: новые инструменты для анализа данных SNP по всему геному. Биоинформатика 27 (21): 3070–3071
CAS Статья PubMed PubMed Central Google Scholar
Khanlou KM, Vandepitte K, Asl LK, Van Bockstaele E (2011) На пути к оптимальной стратегии отбора проб для оценки генетической изменчивости внутри и среди сортов клевера белого ( Trifolium repens L.) с использованием AFLP. Genet Mol Biol 34 (2): 252–258
Статья PubMed PubMed Central Google Scholar
Кишор Г., Гупта С., Пандей А. (2012) Оценка популяционного генетического разнообразия Fagopyrum tataricum с использованием молекулярного маркера SSR.Biochem Syst Ecol 43: 32–41
CAS Статья Google Scholar
Louette D, Charrier A, Berthaud J (1997) Сохранение кукурузы in situ в Мексике: генетическое разнообразие и управление семенами кукурузы в традиционном сообществе. Econ Bot 51 (1): 20–38
Статья Google Scholar
Mantel N (1967) Выявление кластеризации болезней и метод обобщенной регрессии.Cancer Res 27 (2 Часть 1): 209–220
CAS PubMed Google Scholar
Мюррей М.Г., Томпсон В.Ф. (1980) Быстрое выделение высокомолекулярной растительной ДНК. Nucleic Acids Res 8 (19): 4321–4325
CAS Статья PubMed PubMed Central Google Scholar
Нагараджан Л., Смейл М. (2007) Деревенские семенные системы и биологическое разнообразие проса в маргинальных средах Индии.Euphytica 155 (1-2): 167–182
Статья Google Scholar
Нельсон М.Ф., Андерсон Н.О. (2013) Сколько маркерных локусов необходимо? Анализ наборов данных о доминантных маркерах с использованием двух популярных популяционных генетических алгоритмов. Ecol Evol 3 (10): 3455–3470.
PubMed PubMed Central Google Scholar
Охниши О. (1998) Поиск дикого предка гречихи III.Дикий предок культурной гречки обыкновенной и татарской гречихи. Econ Bot 52 (2): 123–133
Статья Google Scholar
Peakall R, Smouse PE (2006) GENALEX 6: генетический анализ в Excel. Популяционно-генетическое программное обеспечение для обучения и исследований. Mol Ecol Notes 6 (1): 288–295
Статья Google Scholar
Peakall R, Smouse PE (2012) GenAlEx 6.5: генетический анализ в Excel.Программное обеспечение популяционной генетики для обучения и исследований — обновление. Биоинформатика 28 (19): 2537–2539
CAS Статья PubMed PubMed Central Google Scholar
Porath J, Flodin P (1959) Гель-фильтрация: метод обессоливания и группового разделения. Nature 183 (4676): 1657–1659
CAS Статья PubMed Google Scholar
Притчард Дж. К., Стивенс М., Доннелли П. (2000) Вывод структуры популяции с использованием данных мультилокусного генотипа.Генетика 155 (2): 945–959
CAS PubMed PubMed Central Google Scholar
Radanielina T, Ramanantsoanirina A, Raboin LM, Frouin J, Perrier X, Brabant P, Ahmadi N (2013) Оригинальные особенности генетического разнообразия риса ( Oryza sativa L.) и важность внутрисортового разнообразия в высокогорье Мадагаскара — убедительные аргументы в пользу сохранения in situ. Genet Resour Crop Evol 60 (1): 311–323
Статья Google Scholar
R Core Team (2013) R: язык и среда для статистических вычислений.R Фонд статистических вычислений, Вена. http://www.r-project.org/
Розенберг Н.А. (2003) DISTRUCT: программа для графического отображения структуры населения. Mol Ecol Notes 4 (1): 137–138
Статья Google Scholar
Руфа Л. (2004) Разработка и использование ресурсов винной гречихи. В: Материалы 9-го международного симпозиума по гречихе, Прага, стр. 252–258
Самберг Л.Х., Фишман Л., Аллендорф Ф.В. (2013) Генетическая структура населения в социальном ландшафте: ячмень в традиционной эфиопской сельскохозяйственной системе.Evol Appl 6 (8): 1133–1145
Статья PubMed PubMed Central Google Scholar
Saunders M (2010) Теряя почву под ногами: неопределенное будущее выращивания гречихи в центре ее происхождения. В кн .: Зотиков В., Парахин Н. (ред.) Успехи исследований гречихи: материалы 11-го международного симпозиума по гречке. ВНИИ зернобобовых и крупяных культур, Орел, с. 60–68
Google Scholar
Сондерс Булан М., Познер Дж. Л., Пенг Д., Эмшвиллер Э, Ван Х, Ли Дж и др. (2015) Старый урожай, новое общество: сохранение и изменение татарского выращивания гречихи в Юньнани, Китай.Human Ecol (в редакции)
Сингх М., Чабейн К., Валкоун Дж., Блейк Т. (2006) Оптимальный размер выборки для оценки генетического разнообразия дикой пшеницы с использованием маркеров AFLP. Genet Resour Crop Evol 53 (1): 23–33
CAS Статья Google Scholar
Скеллам Дж. Г. (1948) Распределение вероятностей, полученное из биномиального распределения путем рассмотрения вероятности успеха как переменной между наборами испытаний. J R Stat Soc Series B Methodol 10 (2): 257–261
Google Scholar
Tsuji K, Ohnishi O (2000) Происхождение культурной татарской гречихи ( Fagopyrum tataricum Gaertn.), выявленные анализом RAPD. Genet Resour Crop Evol 47 (4): 431–438
Статья Google Scholar
Vekemans X (2002) AFLP-SURV версия 1.0. Распространяется автором. Laboratoire de Génétique et Ecologie Végétale, Université Libre de Bruxelles, Brussells
Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes M, Frijters A, Pot J, Peleman J, Kuiper M ( ) AFLP: новый метод снятия отпечатков пальцев ДНК.Nucleic Acids Res 23 (21): 4407–4414
CAS. Статья PubMed PubMed Central Google Scholar
Wickham H (2009) ggplot2: элегантная графика для анализа данных [Интернет]. Спрингер, Нью-Йорк. http://had.co.nz/ggplot2/book
Wright S (1921) Системы вязки. Генетика 6: 111–178
CAS PubMed PubMed Central Google Scholar
Yoshida M, Matsudo S, Hagiwara M, Matano T.